Publications
Activin A
Paloviita P, Hautala E, Granskog R et al.
Hsa-miR-92a-3p regulates cell-cycle and signaling programs during human extra-embryonic lineage commitment
From the lab of Sanna Vuoristo, University of Helsinki and Helsinki University Hospital, Finland
Kaufmann C, Sting S, Dai C et al.
Comprehensive CRISPR/Cas9-based mutagenesis identifies single-amino acid substitutions that abrogate SPEN function in X inactivation
From the lab of Anton Wutz, Institute of Molecular Health Sciences, Department of Biology, ETH Zurich, Switzerland
Sozza F, Romano A, D’Elia N et al.
Combinatorial and Inducible CRISPRa/i Enables Canalized hiPSC Forward Programming and Iterative Refinement via Single-Cell Genomics
From the lab of Alessandro Bertero, Department of Molecular Biotechnology and Health Sciences, University of Turin, Italy
Used:
- Recombinant human NT-3 protein (Qk058)
- Recombinant human activin A protein (Qk001-CTG)
- Recombinant human activin A protein (Qk001)
- Recombinant human BDNF protein (Qk050-CTG)
- Recombinant human BDNF protein (Qk050)
- Recombinant human FGF-2 (145 aa) protein (Qk025-CTG)
- Recombinant human FGF-2 (145 aa) protein (Qk025)
- Recombinant human BMP-4 protein (Qk038)
Fiancette RB, Huang J, Stephens C et al.
High-purity stem cell-derived β-cells recapitulate key transcriptional and functional features of human islets
From the lab of Dr İldem Akerman, University of Birmingham, UK
Kafkia E, Pladevall-Morera D, Argemi-Muntadas L et al.
TCA cycle rewiring underpins histone acetylation sourcing and cell-fate transitions during exit from naive pluripotency
From the lab of Jan Jakub Zylicz, Novo Nordisk Foundation Center for Stem Cell Medicine—reNEW, University of Copenhagen, Denmark
Hernanz M, Matas D, Mirasierra M et al.
Intramuscular Transplantation of Human Pluripotent Stem Cell-derived Pancreatic Endocrine Cells in Mice
DOI: DOI: 10.3791/69562
From the lab of Alberto Bartolomé, Biomedical Research Institute, Madrid
Rossignoli G, Oberhuemer M, Brun IS et al.
Serum coating enables feeder-free culture of naive human pluripotent stem cells preserving developmental potential
From the labs of Jan Jakub Żylicz (Novo Nordisk Foundation Center for Stem Cell Medicine, University of Copenhagen, Denmark), Martin Leeb (Max Perutz Labs, University of Vienna, Austria) and Graziano Martello (University of Padua, Italy)
Used:
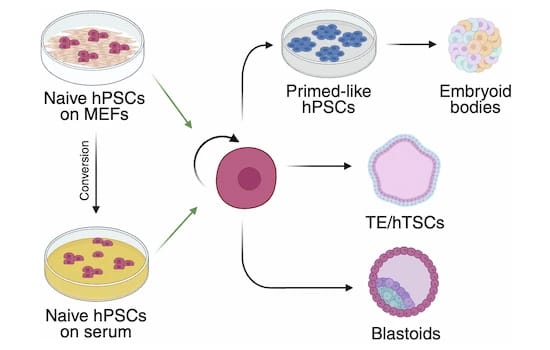
This publication describes a feeder cell-free culture system based on serum coating that supports long-term maintenance of naive human pluripotent stem cells (hPSCs).
Naive hPSC represent a preimplantation epiblast state able to efficiently differentiate into embryonic and extraembryonic pre-implantation lineages. Their maintenance routinely relies on co-culture with mouse embryonic fibroblast (MEFs) as feeder cells, a method prone to variability.
In a collaborative effort, five independent laboratories tested thirty serum batches for the culture of eight naïve hPSCs lines. Cells cultured on serum coating displayed growth kinetics, clonogenic capacity, mutation rates, and global gene expression profiles comparable to MEF-based cultures. Naive hPSCs efficiently underwent germ layer specification, retained trophectoderm competence, and generated blastoids with efficiency similar to MEF-based cultures.
This study showed that serum coating provides a scalable, cost-effective, and robust alternative to feeder-based systems, facilitating larger-scale applications of naive hPSCs and enabling more reproducible mechanistic studies.
This study used Qkine animal origin-free proteins:
Bottini S (Thesis)
Animal proteins from stem cells as an alternative to reduce the ecological and climate impact of animal farming
DOI: Thesis
Used:
- Recombinant bovine/porcine FGF-2 (145 aa) protein (Qk040-FG)
- Recombinant bovine/porcine FGF-2 (145 aa) protein (Qk040)
- Recombinant human TGF-β1 PLUS™ protein (Qk010-CTG)
- Recombinant human TGF-β1 PLUS™ protein (Qk010)
- Recombinant FGF2-G3 (145 aa) protein (Qk052)
- Recombinant human TGF-β3 protein (Qk054-CTG)
- Recombinant human TGF-β3 protein (Qk054)
- Recombinant human NRG-1 protein (Qk045)
- Recombinant human activin A protein (Qk001-CTG)
- Recombinant human activin A protein (Qk001)
- Recombinant human BMP-4 protein (Qk038)
Zamarian V, Monaco L, Marras M et al.
Gene Expression at the Pluripotency Stage Predicts Pancreatic Endocrine Differentiation in iPSC Clones
From the lab of Valeria Sordi, IRCCS Ospedale San Raffaele
Skowronska-Krawczyk et al.
Age-Driven Lipid Remodeling Activates Lysosome-Mediated Plasma Membrane Repair
From the lab of Lauren Albrecht, University of California, Los Angeles
Keskinen T, Jalil S, Gümüşoğlu I et al.
Genetic Correction of the Most Common Mutation Causing Primary Hyperoxaluria Restores Enzyme Localization and Oxalate Metabolism
From the lab of Mervi E. Hyvönen, University of Helsinki, Finland
Azami T, Patton EE and Nichols J.
Simplified In Vitro Generation of Human Gastruloids for Modelling Early Development
From the lab of Jennifer Nichols Institute of Genetics and Cancer, University of Edinburgh, UK
Becca S, Bianchi S, Hahn EM et al.
Opposing CTCF and GATA4 activities set the pace of chromatin topology remodeling during cardiomyogenesis
From the lab of Alessandro Bertero, University of Turin, Italy
Chen C, Wu J, Wang X et al.
Signaling reprogramming via Stat3 activation unravels high-fidelity human post-implantation embryo modeling
From the lab of José Silva, Guangzhou National Laboratory, Guangdong, China
Bernardo, Edgar et al.
HNF1A and A1CF coordinate a beta cell transcription-splicing axis that is disrupted in type 2 diabetes
From the lab of Jorge Ferrer, Centre for Genomic Regulation, the Barcelona Institute of Science and Technology, Spain
Van Nerum, K., Wenzel, A., Argemi-Muntadas, L. et al.
α-Ketoglutarate promotes trophectoderm induction and maturation from naive human embryonic stem cells
From the lab of Jan Jakub Zylicz, University of Copenhagen
Garitta E
Modelling Cholestasis in vitro Using Hepatocytes Derived from Human Induced Pluripotent Stem Cells
Huang, T et al.
Inhibition of PRC2 enables self-renewal of blastoid-competent naive pluripotent stem cells from chimpanzee
From the lab of Hideki Masaki, Institute of Science, Tokyo
Zorzan I, Pellegrini M, Arboit M et al.
The transcriptional regulator ZNF398 mediates pluripotency and epithelial character downstream of TGF-beta in human PSCs
From the lab of Graziano Martello, University of Padua
Beucher A, Miguel-Escalada I, Balboa D et al.
The HASTER lncRNA promoter is a cis-acting transcriptional stabilizer of HNF1A
From the lab of Jorge Ferrer, Centre for Genomic Regulation (CRG)
Bao M, Cornwall-Scoones J, Sanchez-Vasquez E et al.
Stem cell-derived synthetic embryos self-assemble by exploiting cadherin codes and cortical tension
From the lab of Magdalena Zernicka-Goetz, University of Cambridge
Meek S, Watson T, Eory L et al.
Stem cell-derived porcine macrophages as a new platform for studying host-pathogen interactions
From the lab of Tom Burdon, University of Edinburgh
Azami T, Theeuwes B, Ton M-LN et al.
STAT3 signalling enhances tissue expansion during postimplantation mouse development
From the lab of Dr Jennifer Nichols, University of Cambridge
Balmas E, Ratto ML, Snijders KE et al.
Single Cell Transcriptional Perturbome in Pluripotent Stem Cell Models
From the lab of Alessandro Bertero, University of Turin
Guo M, Wu J, Chen C et al.
Self-renewing human naïve pluripotent stem cells dedifferentiate in 3D culture and form blastoids spontaneously
From the lab of José Silva, Guangzhou Laboratory
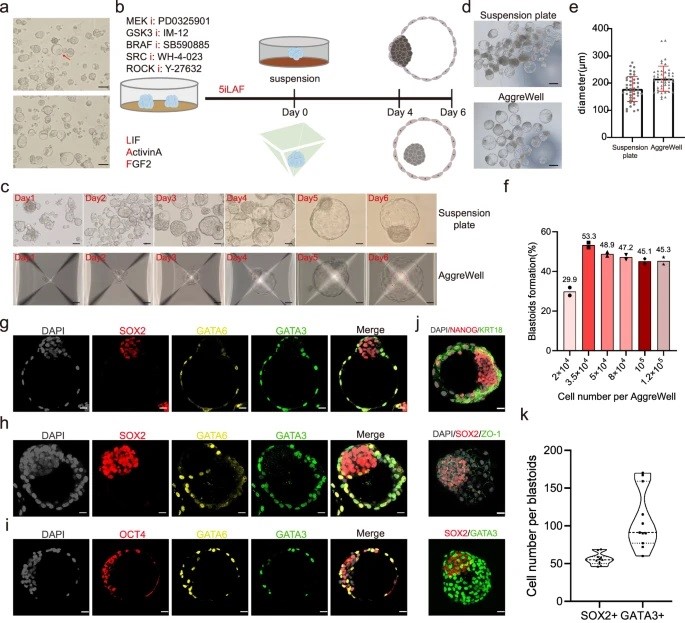
A huge challenge in understanding human early embryo cell fate is due to limited access and ethical concerns. Recent research, however, from José C. R. Silva’s lab at Guangzhou National Laboratory, drawing from single-cell sequencing, suggests a conserved lineage specification process between human and mouse embryos. Blastoids, emerging models for early embryo development, generated solely from hnPSCs, offer insights into blastocyst formation without altering culture conditions. Self-renewing human naïve pluripotent stem cells (hnPSCs) spontaneously form blastoids in 3D culture, mimicking early human blastocysts. This process, mediated by the GSK3 inhibitor IM-12 in 5iLAF medium, involves upregulation of oxidative phosphorylation genes. hnPSCs dedifferentiate into E5 embryo-like intermediates, expressing SOX2/OCT4 and GATA6, which specify trophoblast fate by day 3, coinciding with blastoid formation. This was a fantastic paper to read as it is clear how this spontaneous blastoid formation highlights the importance of culture conditions and provides a new platform to study human embryo development in vitro, potentially reshaping our understanding of hnPSCs and embryo development.
Farbergshagen, AC (Thesis)
Role of mechanotransduction in pancreatic endocrine cell fate acquisition in SC-islets
DOI: Thesis
Truszkowski L, Bottini S, Bianchi S et al.
Refined and benchmarked homemade media for cost-effective, weekend-free human pluripotent stem cell culture
From the lab of Alessandro Bertero, University of Turin in collaboration with Qkine
Used:
- Recombinant human activin A protein (Qk001)
- Recombinant human FGF-2 (145 aa) protein (Qk025)
- Recombinant human FGF-2 (154 aa) protein (Qk027)
- Recombinant human BMP-4 protein (Qk038)
- Recombinant human NRG-1 protein (Qk045)
- Recombinant FGF2-G3 (145 aa) protein (Qk052)
- Recombinant FGF2-G3 (154 aa) protein (Qk053)
- Recombinant human TGF-β3 protein (Qk054)
- Recombinant human TGF-β1 PLUS™ protein (Qk010)
- Thermostable FGF-2 discovery kit (Qk502)
Cell therapy is becoming a possibility for many previously untreatable conditions, and it should be accessible to everyone. Creating a cost-effective, reliable and reproducible way of culturing human induced pluripotent stem cells (hiPSCs) in a range of research labs, and allowing large scale culture for gene-editing purposes takes us one step closer to this.
Using high potency thermostable Qkine 145 amino acid FGF-G3 reduce FGF-2 use 8-fold and for weekend-free culture reduced media use by 57%. This makes hiPSCs a more accessible model for many labs doing basic and translational research.
Militi S, Nibhani R, Jalali M and Pauklin S.
RBL2-E2F-GCN5 guide cell fate decisions during tissue specification by regulating cell-cycle-dependent fluctuations of non-cell-autonomous signaling
From the lab of Siim Pauklin, University of Oxford
Rosa VS, Sato N and Shahbazi MN et al.
Protocol for generating a 3D culture of epiblast stem cells
From the lab of Dr Marta Shahbazi, MRC Laboratory of Molecular Biology, Cambridge
Drozd AM, Mariani L, Guo X, Goitea V, Menezes NA and Ferretti E.
Progesterone Receptor Modulates Extraembryonic Mesoderm and Cardiac progenitor Specification during Mouse Gastrulation
Weatherbee BAT, Gantner CW, Iwamoto-Stohl LK et al.
Pluripotent stem cell-derived model of the post-implantation human embryo
From the lab of Magdalena Zernicka-Goetz, University of Cambridge
Miguel-Escalada I, Maestro MÁ, Balboa D et al.
Pancreas agenesis mutations distrupt a lead enhancer controlling a developmental enhancer cluster
From the lab of Jorge Ferrer, Centre for Genomic Regulation (CRG)
Luo L, Foster NC, Man KL et al.
Hydrostatic pressure promotes chondrogenic differentiation and microvesicle release from human embryonic and bone marrow stem cells
From the lab of Alicia El Haj, University of Birmingham
Jalil S, Keskinen T, Juutila J et al.
Genetic and functional correction of argininosuccinate lyase deficiency using CRISPR adenine base editors
From the lab of Kirmo Wartiovaara, University of Helsinki
Darrigrand J-F, Isaacson A and Spagnoli FM
Generation of human iPSC-derived pancreatic organoids to study pancreas development and disease
Rossignoli G, Oberhuemer M, Brun IS et al.
Feeder-free culture of naive human pluripotent stem cells retaining embryonic, extraembryonic and blastoid generation potential
From the lab of Graziano Martello, University of Padua
Farhan F, Trivedi M, Di Wu P et al.
Extracellular matrices modulate differentiation of human embryonic stem cell-derived hepatocyte-like cells with spatial hepatic features
From the lab of Wei Cui, Imperial College London
Gang N
Exploring the link between dioxin exposure and diabetes risk: contributions of the islet aryl hydrocarbon receptor
DOI: Thesis
Williams TL, Macrae RGC, Kuc RE, Brown AJH, Maguire JJ, Davenport AP.
Expanding the apelin receptor pharmacological toolbox using novel fluorescent ligands
From the lab of Anthony Davenport, University of Cambridge
Carbognin E, Carlini V, Panariello F et al.
Esrrb guides naive pluripotent cells through the formative transcriptional programme
From the labs of Jamie A. Hackett, European Molecular Biology Laboratory EMBL-Rome, Davide Cacchiarelli, Telethon Institute of Genetics and Medicine and Graziano Martello, University of Padua.
Jobbins AM, Haberman N, Artigas N et al.
Dysregulated RNA polyadenylation contributes to metabolic impairment in non-alcoholic fatty liver disease
From the lab of Santiago Vernia, LMS London Institute of Medical Sciences
Weatherbee BAT, Weberling A, Gantner CW et al.
Distinct pathways drive anterior hypoblast specification in the implanting human embryo
From the lab of Magdalena Zernicka-Goetz, University of Cambridge

It has been a pleasure to read about the intricate dance of signaling coordination between the epiblast, trophectoderm, and hypoblast during early human embryonic development in this recent publication from the lab of Magdalena Zernicka-Goetz at the California Institute of Technology/University of Cambridge. Using human embryos and stem cell models, the research reveals NODAL dependency in anterior hypoblast specification, contrasting roles of BMP in mouse and human anterior signaling center maintenance, and the importance of NOTCH signaling in human epiblast survival. Comparative analysis highlights conserved and species-specific factors driving embryonic development. Specifically, NODAL, BMP, and NOTCH play crucial roles in anterior hypoblast formation, with signaling dynamics changing significantly post-implantation. The fantastic research underscores the complexity of signaling pathways during implantation and emphasizes the importance of further investigations to elucidate their roles comprehensively. These findings clearly contribute to understanding early human embryonic development and provide insights for improving stem cell-derived embryo-like models.
Used human activin A (Qk001) at 25 ng/ml for the culture of human embryos.
Ragusa D, Suen C-W, Cortés GT et al.
Dissecting infant leukemia developmental origins with a hemogenic gastruloid model
From the lab of Cristina Pina, Brunel University.
Used recombinant human activin A PLUS™ protein (Qk005) at 100 ng/ml for mouse hemogenic gastruloid assembly.
Barsby T, Ibrahim H, Lithovius V et al.
Differentiating functional human islet-like aggregates from pluripotent stem cells
From the lab of Timo Otonkoski, University of Helsinki
Used human activin A (Qk001) at 100 ng/ml and 10 ng/ml for iPSC differentiation into definitive endoderm.
Boikova A, Quaife-Ryan GA, Batho CAP et al.
A transient modified mRNA encoding Myc and Cyclin T1 induces cardiac regeneration and improves cardiac function after myocardial injury
From the labs of Catherine H. Wilson, University of Cambridge and James E. Hudson, QIMR Berghofer Medical Research Institute
Used recombinant activin A (Qk001) at 50 ng/ml and zebrafish FGF-2 (Qk002) at 20 ng/ml for human mesoderm induction.
Dias A, Pascual-Mas P, Torregrosa-Cortés G et al.
Opposing Nodal and Wnt signalling activities govern the emergence of the mammalian body plan
From the lab of Alfonso Martinez Arias, Universitat Pompeu Fabra
Used recombinant human activin A PLUS™ protein (Qk005) at 25 ng/ml or 100 ng/ml for mouse gastruloid culture.
Frenster JD, Babin S, Casani-Galdon P et al.
Mosaic gastruloids reveal a temporal restriction for developmental cell competition
From the lab of A. Martinez Arias, University Pompeu Fabra
Used recombinant human activin A PLUS™ protein (Qk005) at 25 ng/ml or 100 ng/ml for gastruloid culture and recombinant mouse LIF protein (Qk018) at 10 ng/ml for mouse embryonic stem cell culture.
Kinoshita M, Barber M, Mansfield W et al.
Capture of Mouse and Human Stem Cells with Features of Formative Pluripotency
From the lab of Austin Smith, University of Cambridge & University of Exeter.
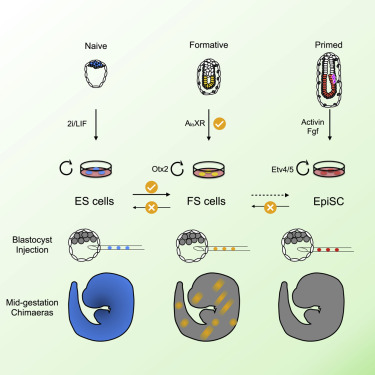
In the study of embryonic stem cells, stem cells representative of naïve and primed pluripotency have been well established in the forms of embryonic stem cells (ESCs) and epiblast-derived stem cells (EpiSCs). In this study Kinoshita et al. fill the gap between early and late pluripotency in describing an intermediate state; formative stem (FS) cells. FS cells differ from both ESCs and EpiSCs, a difference beautifully exemplified by their relative contribution to chimeras. Compared with ESCs, which readily contribute to chimeras, FS chimera contribution is less frequent, and their contribution is less evenly distributed. EpiSCs on the other hand do not generally contribute to chimeras at all. FS cells were established by culturing E5.5 epiblasts, or ES cells, in N2B27 media supplemented with a low dose of Qkine Activin A alongside a Wnt inhibitor and pan-retinoic acid receptor inverse agonist. We are proud our growth factors could be part of such an exciting finding!
Masaki Kinoshita, first author, MRC Cambridge Stem Cell Institute, University of Cambridge, says:
“Formative” pluripotency exists transiently in early development and naive mouse ES cell differentiation, which cells directly respond to differentiation signals. This paper showed that formative pluripotency is now captured in culture and expands its knowledge including chimaera competency of early embryonic cells.
Wamaitha SE, Grybel KJ, Alanis-Lobato G et al.
IGF1-mediated human embryonic stem cell self-renewal recapitulates the embryonic niche
From the lab of Harry Leitch, Imperial College London
Used recombinant human activin A (Qk001) at 50 ng/ml for embryonic stem cell culture.
Stuart HT et al.
Distinct Molecular Trajectories Converge to Induce Naive Pluripotency
From the lab of José Silva, University of Cambridge
Blackford SJI et al.
Validation of Current Good Manufacturing Practice Compliant Human Pluripotent Stem Cell-Derived Hepatocytes for Cell-Based Therapy
From the lab of Tamir Rashid, Kings College London
Used recombinant human activin A protein (Qk001) at 100 ng/ml and 50 ng/ml for hepatocyte differentiation.
Andreasson L, Evenbratt H, Mobini R and Simonsson S et al.
Differentiation of induced pluripotent stem cells into definitive endoderm on Activin A-functionalized gradient surfaces
From the lab of Stina Simonsson, University of Gothenburg
In embryonic development, growth factors are delivered in a highly controlled and targeted manner, however when differentiating iPSCs the real challenge is to effectively mimic these conditions. Consequently, iPSC differentiation is plagued by issues such as low efficiency and a lack of homogeneity. In their recent paper Andreasson et al. take a step towards improving the differentiation of iPSCs to definitive endoderm. The group employs gold nanoparticles to generate a gradient of immobilised Activin A – a member of the TGF-β superfamily that plays a key role in definitive endoderm development. Using this gradient, the group was able to deliver Activin A in a controlled and localised manor, resulting in more efficient differentiation. By deploying their innovative approach, the group observed a dose dependent response of the cells to Activin A, as defined by expression of differentiation markers SOX17 and GATA4. Their results indicate that it may be possible to define an optimal density of Activin A for definitive endoderm differentiation – a finding that could improve the homogeneity and speed of differentiation. This innovative study is a wonderful example of how reconsidering the way in which growth factors are delivered can lead to advances in our understanding of the precise control of stem cell differentiation and how these cells undertake their fate decisions.
BDNF
Sozza F, Romano A, D’Elia N et al.
Combinatorial and Inducible CRISPRa/i Enables Canalized hiPSC Forward Programming and Iterative Refinement via Single-Cell Genomics
From the lab of Alessandro Bertero, Department of Molecular Biotechnology and Health Sciences, University of Turin, Italy
Used:
- Recombinant human NT-3 protein (Qk058)
- Recombinant human activin A protein (Qk001-CTG)
- Recombinant human activin A protein (Qk001)
- Recombinant human BDNF protein (Qk050-CTG)
- Recombinant human BDNF protein (Qk050)
- Recombinant human FGF-2 (145 aa) protein (Qk025-CTG)
- Recombinant human FGF-2 (145 aa) protein (Qk025)
- Recombinant human BMP-4 protein (Qk038)
Grass T, Cosacak IM, Ordureau A et al.
Isogenic cortical organoids enable precision targeting of APP variant-specific pathways in Alzheimer’s disease
From the lab of Natalia Rodriguez-Muela, DZNE
Used:
Alzheimer’s disease is one of the most prevalent neurodegenerative diseases and the leading cause of dementia, the 7th leading cause of death globally. Currently, there is a lack of suitable and relevant disease models, leading to difficulties in both understanding and treating the disease.
Whilst Alzheimer’s disease primarily affects older adults, early onset familial disease has been shown to be caused by pathogenic mutations in the amyloid precursor protein (APP) gene, these mutations may allow the creation of disease models and therefore a method of testing and developing treatments.
This week’s #Fridayread from the lab of Dr. Natalia Rodriguez-Muela describes the generation of an isogenic panel of hiPSC-derived cortical organoids carrying familial Alzheimer’s disease-associated APP variants or the protective A673T variant. Through the panel of 3D models they identified distinct pathogenic pathways specific to each variant.
Proteomic analysis of the organoids allowed them to determine variant-specific defects which related to proteins dysregulated in post-mortem diseased brains, demonstrating the reliability of their in vitro model. Through this they were able to employ targeted interventions and restore neuronal survival in a variant-specific manner.
This study used Qkine BDNF (Qk050) and GDNF (Qk051) at 20 ng/ml in the differentiation of hiPSCs into cortical spheres.
Featherby SJ, Faulkner EC, Gordon A and Ettelaie C.
Procoagulant Extracellular Vesicles Increase Neuronal Tau expression, Metabolism and Processing Through Tissue Factor and Protease Activated Receptor 2
From the lab of Camille Ettelaie, Hull-York Medical School, UK
Thomas J (Thesis)
SMN Protein Heterogeneity Drives Intrinsic Motor Neuron Vulnerability through Survival and Activity Related Pathways
DOI: Thesis
Used BDNF (Qk050) and GDNF (Qk051) at 10 ng/ml for motor neuron differentiation from iPS.
Macarelli V, Sroka TJ, Hansen JN, Axelsson U, Mick DU and Merkle FT
Proximity proteomics of primary cilia in human hypothalamic neurons
From the lab of Florian T. Merkle, University of Cambridge, UK
Magarotto M, Gawne RT, Vilkaite G, Mason AS, Chen H-J
Familial ALS/FTD-associated RNA-binding deficient TDP-43 mutants cause neuronal and synaptic transcript dysregulation in vitro
From the lab of Han-Jou Chen, University of York
Nuhu-Soso L, Denton H, Goffin DL, Hahn I and Evans GJO.
Neuronal differentiation and activity drive nucleocytoplasmic shuttling of the intellectual disability kinase TLK2
From the lab of Gareth J O Evans University of York
Buchner F, Dokuzluoglu Z, Thomas J et al.
Synchronous 3D patterning of diverse CNS progenitors generates motor neurons of broad axial identity
From the lab of Dr. Natalia Rodriguez-Muela, German Center for Neurodegenerative Diseases e.V. (DZNE)
Chen HJC, Yang A, Mazzaferro S et al.
Profiling human hypothalamic neurons reveals a candidate combination drug therapy for weight loss
From the labs of John C. Marioni, European Bioinformatics Institute and Florian T. Merkle, University of Cambridge
Page T, Musi CA, Bakker SE et al.
Parkinson’s associated protein DJ-1 regulates intercellular communication via extracellular vesicles in oxidative stress
From the lab of Mariaelena Repici, Aston University
Used:

This publication explores disease mechanisms in an autosomal recessive form of Parkinson’s disease caused by mutations in the DJ-1 protein. Parkinson’s disease is neurodegenerative disorder effecting the central and peripheral nervous system, it is common and effects 1% of people over 60. Parkinson’s is characterized by the loss of dopaminergic neurons and accumulation of Lewy bodies in the substantia nigra area of the brain.
DJ-1 protein, encoded by the Park7 gene, is a multifunctional protein which has been shown to have roles in the regulation of apoptosis ad the immune system and also dopamine synthesis and uptake. This study looks at the role of DJ-1 in extracellular vesicles, which are secreted by brain cells and regulate synaptic function and neural development.
Proteomic analysis of extracellular vesicles from DJ-1 deficient cells showed a distinct protein expression profile. Challenging DJ-1 KO cells with rotenone, a naturally occurring pesticide which induces oxidative stress, showed DJ-1 deficient cells have reduced numbers of extracellular vesicles and increased macrophage migration.
These result indicate DJ-1 regulates the number, cargo and functional effects of extracellular vesicles released from neuronal cells upon oxidative stress, offering new insights into disease progression and novel therapeutic targets.
Pivoňková H, Sitnikov S, Kamen Y et al.
Heterogeneity in oligodendrocyte precursor cell proliferation is dynamic and driven by passive bioelectrical properties
From the lab of Ragnhildur Thóra Káradóttir, Cambridge Stem Cell Institute
Macarelli V, Harding EC, Gershlick DC, Merkle FT.
A short sequence targets transmembrane proteins to primary cilia
From the lab of Florian Merkle, University of Cambridge
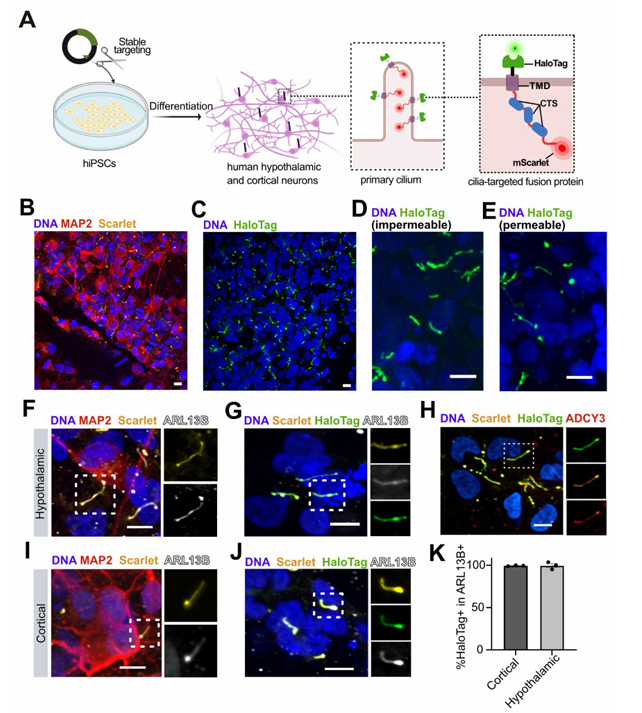
Macarelli et al demonstrate the importance of using the right cells and tools for the study of neural stem cell models in this recent publication. They explored the use of different ciliary targeting sequences to fluorescently label and measure cilia. This allows the study of the structure and function of these important sensory and signaling structures in differentiated neural cells. An exciting addition to the knowledge of tools to study neural differentiation from the lab of Dr Florian Merkle at the Cambridge Stem Cell Institute.
Used recombinant human BDNF (Qk050) at 10 μg/ml for hypothalamic neuron differentiation from iPSC.
Agarwal D et al.
Human retinal ganglion cell neurons generated by synchronous BMP inhibition and transcription factor mediated reprogramming
From the lab of Karl Wahlin, University of California San Diego
Used recombinant human BDNF protein (Qk050) at 50 ng/ml and recombinant human GDNF protein (Qk051) at 10 ng/ml for differentiation of PSCs in retinal ganglion cell induced neurons.
Agarwal D & Wahlin K
Differentiation of RGC Induced Neurons (RGC-iNs)
From the lab of Karl Wahlin, University of California San Diego
BMP-2
Holthaus D, Le HD, Matzner L et al.
Organoids serve as viable in vitro model for functional precision medicine for mesonephric-like adenocarcinoma of the ovary
From the lab of Dr Nina Hedemann, Kiel University, Germany
Huang TC, Wang YF, Vazquez-Ferrer E et al.
Sex-specific chromatin remodelling safeguards transcription in germ cells
From the lab of Petra Hajkova, MRC London Institute of Medical Sciences
Luo L, Foster NC, Man KL et al.
Hydrostatic pressure promotes chondrogenic differentiation and microvesicle release from human embryonic and bone marrow stem cells
From the lab of Alicia El Haj, University of Birmingham
Stucchi S, Sepulveda-Rincon LP, Dion C et al.
High resolution multi-scale profiling of embryonic germ cell-like cells derivation reveals pluripotent state transitions in humans
Yimiti D, Uchibe K, Toriyama M et al.
CD1530, selective RARγ agonist, facilitates Achilles tendon healing by modulating the healing environment including less chondrification in a mouse model
From the lab of Shigeru Miyaki, Hiroshima University
BMP-4
Sozza F, Romano A, D’Elia N et al.
Combinatorial and Inducible CRISPRa/i Enables Canalized hiPSC Forward Programming and Iterative Refinement via Single-Cell Genomics
From the lab of Alessandro Bertero, Department of Molecular Biotechnology and Health Sciences, University of Turin, Italy
Used:
- Recombinant human NT-3 protein (Qk058)
- Recombinant human activin A protein (Qk001-CTG)
- Recombinant human activin A protein (Qk001)
- Recombinant human BDNF protein (Qk050-CTG)
- Recombinant human BDNF protein (Qk050)
- Recombinant human FGF-2 (145 aa) protein (Qk025-CTG)
- Recombinant human FGF-2 (145 aa) protein (Qk025)
- Recombinant human BMP-4 protein (Qk038)
Yamagchi A, Ishikawa K and Akamatsu W
Modeling the pathological brain-gut axis in Parkinson’s disease using human iPSC derived brain-intestinal assembloids
From the lab of Wado Akamatsu, Juntendo University, Tokyo, Japan
Bottini S (Thesis)
Animal proteins from stem cells as an alternative to reduce the ecological and climate impact of animal farming
DOI: Thesis
Used:
- Recombinant bovine/porcine FGF-2 (145 aa) protein (Qk040-FG)
- Recombinant bovine/porcine FGF-2 (145 aa) protein (Qk040)
- Recombinant human TGF-β1 PLUS™ protein (Qk010-CTG)
- Recombinant human TGF-β1 PLUS™ protein (Qk010)
- Recombinant FGF2-G3 (145 aa) protein (Qk052)
- Recombinant human TGF-β3 protein (Qk054-CTG)
- Recombinant human TGF-β3 protein (Qk054)
- Recombinant human NRG-1 protein (Qk045)
- Recombinant human activin A protein (Qk001-CTG)
- Recombinant human activin A protein (Qk001)
- Recombinant human BMP-4 protein (Qk038)
Baxter R (Thesis)
Targeting phosphatidylserine exposure in pro-coagulant platelets
Baxter R, Crosby A, Foster HR et al.
ATP11A and ATP11C are plasma membrane phosphatidylserine flippases in in vitro human megakaryocytes
From the lab of Matthew Harper, University of Cambridge
Used human BMP-4 (Qk038) at 10 ng/ml for mesoderm induction prior to megakaryocyte differentiation.
Becca S, Bianchi S, Hahn EM et al.
Opposing CTCF and GATA4 activities set the pace of chromatin topology remodeling during cardiomyogenesis
From the lab of Alessandro Bertero, University of Turin, Italy
Barbieri E and Chambers I
OTX2 controls chromatin accessibility to direct somatic versus germline differentiation
From the lab of Professor Ian Chambers, The University of Edinburgh
Azami T, Theeuwes B, Ton M-LN et al.
STAT3 signalling enhances tissue expansion during postimplantation mouse development
From the lab of Dr Jennifer Nichols, University of Cambridge
Balmas E, Ratto ML, Snijders KE et al.
Single Cell Transcriptional Perturbome in Pluripotent Stem Cell Models
From the lab of Alessandro Bertero, University of Turin
Truszkowski L, Bottini S, Bianchi S et al.
Refined and benchmarked homemade media for cost-effective, weekend-free human pluripotent stem cell culture
From the lab of Alessandro Bertero, University of Turin in collaboration with Qkine
Used:
- Recombinant human activin A protein (Qk001)
- Recombinant human FGF-2 (145 aa) protein (Qk025)
- Recombinant human FGF-2 (154 aa) protein (Qk027)
- Recombinant human BMP-4 protein (Qk038)
- Recombinant human NRG-1 protein (Qk045)
- Recombinant FGF2-G3 (145 aa) protein (Qk052)
- Recombinant FGF2-G3 (154 aa) protein (Qk053)
- Recombinant human TGF-β3 protein (Qk054)
- Recombinant human TGF-β1 PLUS™ protein (Qk010)
- Thermostable FGF-2 discovery kit (Qk502)
Cell therapy is becoming a possibility for many previously untreatable conditions, and it should be accessible to everyone. Creating a cost-effective, reliable and reproducible way of culturing human induced pluripotent stem cells (hiPSCs) in a range of research labs, and allowing large scale culture for gene-editing purposes takes us one step closer to this.
Using high potency thermostable Qkine 145 amino acid FGF-G3 reduce FGF-2 use 8-fold and for weekend-free culture reduced media use by 57%. This makes hiPSCs a more accessible model for many labs doing basic and translational research.
Drozd AM, Mariani L, Guo X, Goitea V, Menezes NA and Ferretti E.
Progesterone Receptor Modulates Extraembryonic Mesoderm and Cardiac progenitor Specification during Mouse Gastrulation
Tan J, Virtue S, Norris DM et al.
Limited oxygen in standard cell culture alters metabolism and function of differentiated cells
From the lab of Daniel J. Fazakerley, University of Cambridge

We all know our stem cell cultures are sensitive and high maintenance, we feed them, keep them warm to make sure we get the most accurate, reproducible data. The composition of their media is important, we need the right growth factors, but this week’s Friday read highlights that their exposure to oxygen can be an important factor in the differentiation and function of cellular models. In this very interesting read from the MRC Institute of Metabolic Science they have investigated the impact of oxygen diffusion and media volume on hypoxia-related transcriptional changes in stem cell cultures. They found that decreasing the media volume decreased lactate production and HIF1α expression and increased the functionality of adipocyte, hiPSC-derived hepatocytes and hiPSC-derived cardiac organoid cultures.
Stucchi S, Sepulveda-Rincon LP, Dion C et al.
High resolution multi-scale profiling of embryonic germ cell-like cells derivation reveals pluripotent state transitions in humans
EGF
Asadpour A, Ozsoy N, Napier L et al.
Neural Crest-Derived Stem Cell Secretomes and Extracellular Vesicles Disrupt Glioblastoma through Dual-Pathway Inflammatory Rebalancing
From the lab of Darius Widera, Stem Cell Biology and Regenerative Medicine Group, University of Reading, UK
Pantazaka E, Papakonstantinou D, Roumeliotou A et al.
Repurposing Artesunate to Combat Progression and Metastasis via Targeting Circulating Tumor Cells
From the lab of Kallergi Galatea Laboratory of Biochemistry/Metastatic Signaling, University of Patras, Greece
Fiancette RB, Huang J, Stephens C et al.
High-purity stem cell-derived β-cells recapitulate key transcriptional and functional features of human islets
From the lab of Dr İldem Akerman, University of Birmingham, UK
Hernanz M, Matas D, Mirasierra M et al.
Intramuscular Transplantation of Human Pluripotent Stem Cell-derived Pancreatic Endocrine Cells in Mice
DOI: DOI: 10.3791/69562
From the lab of Alberto Bartolomé, Biomedical Research Institute, Madrid
Artioli A, Gasparotto M, Rossetti AC et al.
A defined 2D system for generating and expanding human basal radial glia from iPSCs
From the lab of Julia Ladewig, Central Institute of Mental Health (ZI), Heidelberg University/Medical Faculty Mannheim, Germany
Tran D, Tolley C, Morris T et al.
Recapitulating whipworm development in vitro using caecaloids
From the lab of Dr Maria Duque-Correa, University of Cambridge
Rossignoli G, Oberhuemer M, Brun IS et al.
Serum coating enables feeder-free culture of naive human pluripotent stem cells preserving developmental potential
From the labs of Jan Jakub Żylicz (Novo Nordisk Foundation Center for Stem Cell Medicine, University of Copenhagen, Denmark), Martin Leeb (Max Perutz Labs, University of Vienna, Austria) and Graziano Martello (University of Padua, Italy)
Used:
This publication describes a feeder cell-free culture system based on serum coating that supports long-term maintenance of naive human pluripotent stem cells (hPSCs).
Naive hPSC represent a preimplantation epiblast state able to efficiently differentiate into embryonic and extraembryonic pre-implantation lineages. Their maintenance routinely relies on co-culture with mouse embryonic fibroblast (MEFs) as feeder cells, a method prone to variability.
In a collaborative effort, five independent laboratories tested thirty serum batches for the culture of eight naïve hPSCs lines. Cells cultured on serum coating displayed growth kinetics, clonogenic capacity, mutation rates, and global gene expression profiles comparable to MEF-based cultures. Naive hPSCs efficiently underwent germ layer specification, retained trophectoderm competence, and generated blastoids with efficiency similar to MEF-based cultures.
This study showed that serum coating provides a scalable, cost-effective, and robust alternative to feeder-based systems, facilitating larger-scale applications of naive hPSCs and enabling more reproducible mechanistic studies.
This study used Qkine animal origin-free proteins:
Huang Y, Xu L, Fu H et al.
Tumor suppressor protein p53 governs human trophoblast lineage development
From the lab of Pentao Liu School of Biomedical Sciences, Li Kai Shing Faculty of Medicine, China
Ramage DE, Wieske LHE, Crowe C at al.
LRRC58 defines an E3 ubiquitin ligase complex sensitive to cysteine abundance
From the lab of Richard Timms, Cambridge Institute for Therapeutic Immunology and Infectious Disease, University of Cambridge, UK
Rastovic U, Campinoti S, Wei L et al.
Comprehensive analysis of extracellular matrix remodelling via cyclophilin inhibition in human models of alcohol-related liver fibrosis
From the lab of Elena Palma, Foundation for Liver Research and King’s College Hospital, London, UK
Schwarz LC, Shannon MJ, McNeill G et al.
A blastocyst-derived in vitro model of the human chorion
From the lab Marta N. Shahbazi, MRC Laboratory of Molecular Biology, Cambridge, UK
Van Nerum, K., Wenzel, A., Argemi-Muntadas, L. et al.
α-Ketoglutarate promotes trophectoderm induction and maturation from naive human embryonic stem cells
From the lab of Jan Jakub Zylicz, University of Copenhagen
Christensen JB, Donovan APA, Marzieh Moradi M et al.
A conserved differentiation program facilitates inhibitory neuron production in the developing mouse and human cerebellum
From the lab of Sumru Bayin, Gurdon Institute, University of Cambridge
Darrigrand J-F, Isaacson A and Spagnoli FM
Generation of human iPSC-derived pancreatic organoids to study pancreas development and disease
Rossignoli G, Oberhuemer M, Brun IS et al.
Feeder-free culture of naive human pluripotent stem cells retaining embryonic, extraembryonic and blastoid generation potential
From the lab of Graziano Martello, University of Padua
FGF-10
Holthaus D, Le HD, Matzner L et al.
Organoids serve as viable in vitro model for functional precision medicine for mesonephric-like adenocarcinoma of the ovary
From the lab of Dr Nina Hedemann, Kiel University, Germany
Tran D, Tolley C, Morris T et al.
Recapitulating whipworm development in vitro using caecaloids
From the lab of Dr Maria Duque-Correa, University of Cambridge
Nakashima K, Tanaka J, Matsuno E et al.
Development of a high-efficiency induction system for embryonic oral epithelium from human pluripotent stem cells
From the lab of Kenji Mishima, Showa Medical University, Tokyo, Japan
van Nunen S
Investigating the effects of obesity on kidney tubuloids and toward combining them with liver and adipoids in a multi-organ-on-a-chip model
DOI: Thesis
Matsuno E et al.
Human iPSC-derived salivary gland cell sheets integrate with injured glands to form glandular structures
From the lab of Junichi Tanaka Showa Medical University, Tokyo, Japan
Iyer DP, Khoei HH, van der Weijden VA et al.
mTOR activity paces human blastocyst stage developmental progression
From the lab of Dr. Aydan Bulut-Karslıoğlu, Max Planck Institute for Molecular Genetics
Agarwal R, Dittmar T, Beer HD et al.
Human epidermis organotypic cultures, a reproducible system recapitulating the epidermis in vitro
From the labs of Emmanuel Contassot and Alexander A. Navarini, University of Basel
Darrigrand J-F, Isaacson A and Spagnoli FM
Generation of human iPSC-derived pancreatic organoids to study pancreas development and disease
FGF-2
Clement M, Gibbs A, Begum A et al.
Astrocyte immunosuppressive activity in glioblastoma depends on ZEB1 and is counteracted by CXCL14
From the lab of Florian Siebzehnrubl, Cardiff University School of Biosciences, Cardiff, Wales
Pozo Garcia V, Çobanoğlu T, Riga K et al.
Glycine: The missing link between carbohydrate and xenobiotic metabolism in the maturing human hepatocyte
From the lab of Sofia Moco, Amsterdam Institute of Molecular and Life Sciences (AIMMS), Vrije Universiteit Amsterdam, The Netherlands
Kaufmann C, Sting S, Dai C et al.
Comprehensive CRISPR/Cas9-based mutagenesis identifies single-amino acid substitutions that abrogate SPEN function in X inactivation
From the lab of Anton Wutz, Institute of Molecular Health Sciences, Department of Biology, ETH Zurich, Switzerland
Sozza F, Romano A, D’Elia N et al.
Combinatorial and Inducible CRISPRa/i Enables Canalized hiPSC Forward Programming and Iterative Refinement via Single-Cell Genomics
From the lab of Alessandro Bertero, Department of Molecular Biotechnology and Health Sciences, University of Turin, Italy
Used:
- Recombinant human NT-3 protein (Qk058)
- Recombinant human activin A protein (Qk001-CTG)
- Recombinant human activin A protein (Qk001)
- Recombinant human BDNF protein (Qk050-CTG)
- Recombinant human BDNF protein (Qk050)
- Recombinant human FGF-2 (145 aa) protein (Qk025-CTG)
- Recombinant human FGF-2 (145 aa) protein (Qk025)
- Recombinant human BMP-4 protein (Qk038)
Asadpour A, Ozsoy N, Napier L et al.
Neural Crest-Derived Stem Cell Secretomes and Extracellular Vesicles Disrupt Glioblastoma through Dual-Pathway Inflammatory Rebalancing
From the lab of Darius Widera, Stem Cell Biology and Regenerative Medicine Group, University of Reading, UK
Pantazaka E, Papakonstantinou D, Roumeliotou A et al.
Repurposing Artesunate to Combat Progression and Metastasis via Targeting Circulating Tumor Cells
From the lab of Kallergi Galatea Laboratory of Biochemistry/Metastatic Signaling, University of Patras, Greece
Kafkia E, Pladevall-Morera D, Argemi-Muntadas L et al.
TCA cycle rewiring underpins histone acetylation sourcing and cell-fate transitions during exit from naive pluripotency
From the lab of Jan Jakub Zylicz, Novo Nordisk Foundation Center for Stem Cell Medicine—reNEW, University of Copenhagen, Denmark
Watson TM, Goatley LC, Meek S et al.
Suidae iPSC-derived macrophages as models for investigating susceptibility and resilience to African swine fever virus
From the lab of Christopher Netherton at The Pirbright Institute, Woking, UK and Tom Burdon, The Roslin Institute, University of Edinburgh, UK.
Naylor K, Webb S, Rajesh D and Mee PJ.
Derivation and characterization of an embryonic‑derived muscle progenitor cell line from Atlantic salmon (Salmo salar)
From the lab of Joe Mee, Roslin Technologies
Artioli A, Gasparotto M, Rossetti AC et al.
A defined 2D system for generating and expanding human basal radial glia from iPSCs
From the lab of Julia Ladewig, Central Institute of Mental Health (ZI), Heidelberg University/Medical Faculty Mannheim, Germany
TCW J, Qian L, Pipalia NH et al.
Cholesterol and matrisome pathways dysregulated in astrocytes and microglia
From the lab of Alison M Goate, Department of Genetics and Genomic Sciences, Icahn School of Medicine at Mount Sinai, New York
Lotz S, Goderie S, Tokas N et al.
Sustained levels of FGF2 maintain undifferentiated stem cell cultures with biweekly feeding
From the lab of Christopher A. Fasano Neural Stem Cell Institute, Rensselaer, New York
Bertucci T, Kakarla S, Winkelman MA et al.
Direct differentiation of human pluripotent stem cells into vascular network along with supporting mural cells
From the lab of Guohao Dai, Northeastern University, Boston, Massachusetts
Rossignoli G, Oberhuemer M, Brun IS et al.
Serum coating enables feeder-free culture of naive human pluripotent stem cells preserving developmental potential
From the labs of Jan Jakub Żylicz (Novo Nordisk Foundation Center for Stem Cell Medicine, University of Copenhagen, Denmark), Martin Leeb (Max Perutz Labs, University of Vienna, Austria) and Graziano Martello (University of Padua, Italy)
Used:
This publication describes a feeder cell-free culture system based on serum coating that supports long-term maintenance of naive human pluripotent stem cells (hPSCs).
Naive hPSC represent a preimplantation epiblast state able to efficiently differentiate into embryonic and extraembryonic pre-implantation lineages. Their maintenance routinely relies on co-culture with mouse embryonic fibroblast (MEFs) as feeder cells, a method prone to variability.
In a collaborative effort, five independent laboratories tested thirty serum batches for the culture of eight naïve hPSCs lines. Cells cultured on serum coating displayed growth kinetics, clonogenic capacity, mutation rates, and global gene expression profiles comparable to MEF-based cultures. Naive hPSCs efficiently underwent germ layer specification, retained trophectoderm competence, and generated blastoids with efficiency similar to MEF-based cultures.
This study showed that serum coating provides a scalable, cost-effective, and robust alternative to feeder-based systems, facilitating larger-scale applications of naive hPSCs and enabling more reproducible mechanistic studies.
This study used Qkine animal origin-free proteins:
Brochard V, Galle A and Jouneau A
Commercially available synthetic hydrogels cannot replace Matrigel in promoting polarization and lumenogenesis of 3D ESC aggregates
From the lab of Alice Jouneau, Université Paris-Saclay, France
Maeding N, Kundully DS, Steinhuber A et al.
Deep immune-phenotyping of HLA-homozygous iPS-cardiomyocytes by spectral flow cytometry
From the lab of Dirk Strunk, Paracelsus Medical University, Salzburg, Austria
Bottini S (Thesis)
Animal proteins from stem cells as an alternative to reduce the ecological and climate impact of animal farming
DOI: Thesis
Used:
- Recombinant bovine/porcine FGF-2 (145 aa) protein (Qk040-FG)
- Recombinant bovine/porcine FGF-2 (145 aa) protein (Qk040)
- Recombinant human TGF-β1 PLUS™ protein (Qk010-CTG)
- Recombinant human TGF-β1 PLUS™ protein (Qk010)
- Recombinant FGF2-G3 (145 aa) protein (Qk052)
- Recombinant human TGF-β3 protein (Qk054-CTG)
- Recombinant human TGF-β3 protein (Qk054)
- Recombinant human NRG-1 protein (Qk045)
- Recombinant human activin A protein (Qk001-CTG)
- Recombinant human activin A protein (Qk001)
- Recombinant human BMP-4 protein (Qk038)
Skowronska-Krawczyk et al.
Age-Driven Lipid Remodeling Activates Lysosome-Mediated Plasma Membrane Repair
From the lab of Lauren Albrecht, University of California, Los Angeles
Azami T, Patton EE and Nichols J.
Simplified In Vitro Generation of Human Gastruloids for Modelling Early Development
From the lab of Jennifer Nichols Institute of Genetics and Cancer, University of Edinburgh, UK
Becca S, Bianchi S, Hahn EM et al.
Opposing CTCF and GATA4 activities set the pace of chromatin topology remodeling during cardiomyogenesis
From the lab of Alessandro Bertero, University of Turin, Italy
Ramage DE, Wieske LHE, Crowe C at al.
LRRC58 defines an E3 ubiquitin ligase complex sensitive to cysteine abundance
From the lab of Richard Timms, Cambridge Institute for Therapeutic Immunology and Infectious Disease, University of Cambridge, UK
Naderlinger E, Murphy C, Feifel E, Jennings P, Gstraunthaler G and Wilmes, A
One-Step Differentiation of Human-Induced Pluripotent Stem Cells into Podocytes
From the lab of Anja Wilmes, Vrije Universiteit Amsterdam, The Netherlands
Mulholland KE, Bourguet M, Cheng N et al.
Pervanadate-induced oxidation relieves autoinhibition of SRC protein tyrosine kinase
Christensen JB, Donovan APA, Marzieh Moradi M et al.
A conserved differentiation program facilitates inhibitory neuron production in the developing mouse and human cerebellum
From the lab of Sumru Bayin, Gurdon Institute, University of Cambridge
Wang Y, Siebzehnrubl D, Weller M et al.
Vortioxetine: A Potential Drug for Repurposing for Glioblastoma Treatment via a Microsphere Local Delivery System
From the lab of Ben Newland, Cardiff University, UK
Huang, T et al.
Inhibition of PRC2 enables self-renewal of blastoid-competent naive pluripotent stem cells from chimpanzee
From the lab of Hideki Masaki, Institute of Science, Tokyo
Zorzan I, Pellegrini M, Arboit M et al.
The transcriptional regulator ZNF398 mediates pluripotency and epithelial character downstream of TGF-beta in human PSCs
From the lab of Graziano Martello, University of Padua
Tan, J. (Thesis)
The effects of culture conditions on cell metabolism and function
DOI: Thesis
Buchner F, Dokuzluoglu Z, Thomas J et al.
Synchronous 3D patterning of diverse CNS progenitors generates motor neurons of broad axial identity
From the lab of Dr. Natalia Rodriguez-Muela, German Center for Neurodegenerative Diseases e.V. (DZNE)
Meek S, Watson T, Eory L et al.
Stem cell-derived porcine macrophages as a new platform for studying host-pathogen interactions
From the lab of Tom Burdon, University of Edinburgh
Balmas E, Ratto ML, Snijders KE et al.
Single Cell Transcriptional Perturbome in Pluripotent Stem Cell Models
From the lab of Alessandro Bertero, University of Turin
Truszkowski L, Bottini S, Bianchi S et al.
Refined and benchmarked homemade media for cost-effective, weekend-free human pluripotent stem cell culture
From the lab of Alessandro Bertero, University of Turin in collaboration with Qkine
Used:
- Recombinant human activin A protein (Qk001)
- Recombinant human FGF-2 (145 aa) protein (Qk025)
- Recombinant human FGF-2 (154 aa) protein (Qk027)
- Recombinant human BMP-4 protein (Qk038)
- Recombinant human NRG-1 protein (Qk045)
- Recombinant FGF2-G3 (145 aa) protein (Qk052)
- Recombinant FGF2-G3 (154 aa) protein (Qk053)
- Recombinant human TGF-β3 protein (Qk054)
- Recombinant human TGF-β1 PLUS™ protein (Qk010)
- Thermostable FGF-2 discovery kit (Qk502)
Cell therapy is becoming a possibility for many previously untreatable conditions, and it should be accessible to everyone. Creating a cost-effective, reliable and reproducible way of culturing human induced pluripotent stem cells (hiPSCs) in a range of research labs, and allowing large scale culture for gene-editing purposes takes us one step closer to this.
Using high potency thermostable Qkine 145 amino acid FGF-G3 reduce FGF-2 use 8-fold and for weekend-free culture reduced media use by 57%. This makes hiPSCs a more accessible model for many labs doing basic and translational research.
Rosa VS, Sato N and Shahbazi MN et al.
Protocol for generating a 3D culture of epiblast stem cells
From the lab of Dr Marta Shahbazi, MRC Laboratory of Molecular Biology, Cambridge
Drozd AM, Mariani L, Guo X, Goitea V, Menezes NA and Ferretti E.
Progesterone Receptor Modulates Extraembryonic Mesoderm and Cardiac progenitor Specification during Mouse Gastrulation
Page T, Musi CA, Bakker SE et al.
Parkinson’s associated protein DJ-1 regulates intercellular communication via extracellular vesicles in oxidative stress
From the lab of Mariaelena Repici, Aston University
Used:
This publication explores disease mechanisms in an autosomal recessive form of Parkinson’s disease caused by mutations in the DJ-1 protein. Parkinson’s disease is neurodegenerative disorder effecting the central and peripheral nervous system, it is common and effects 1% of people over 60. Parkinson’s is characterized by the loss of dopaminergic neurons and accumulation of Lewy bodies in the substantia nigra area of the brain.
DJ-1 protein, encoded by the Park7 gene, is a multifunctional protein which has been shown to have roles in the regulation of apoptosis ad the immune system and also dopamine synthesis and uptake. This study looks at the role of DJ-1 in extracellular vesicles, which are secreted by brain cells and regulate synaptic function and neural development.
Proteomic analysis of extracellular vesicles from DJ-1 deficient cells showed a distinct protein expression profile. Challenging DJ-1 KO cells with rotenone, a naturally occurring pesticide which induces oxidative stress, showed DJ-1 deficient cells have reduced numbers of extracellular vesicles and increased macrophage migration.
These result indicate DJ-1 regulates the number, cargo and functional effects of extracellular vesicles released from neuronal cells upon oxidative stress, offering new insights into disease progression and novel therapeutic targets.
Tan J, Virtue S, Norris DM et al.
Oxygen is a critical regulator of cellular metabolism and function in cell culture
Iyer DP, Khoei HH, van der Weijden VA et al.
mTOR activity paces human blastocyst stage developmental progression
From the lab of Dr. Aydan Bulut-Karslıoğlu, Max Planck Institute for Molecular Genetics
Beltran-Rendon C, Price CJ, Glen K et al.
Modeling the selective growth advantage of genetically variant human pluripotent stem cells to identify opportunities for manufacturing process control
From the lab of Robert Thomas, Loughborough University
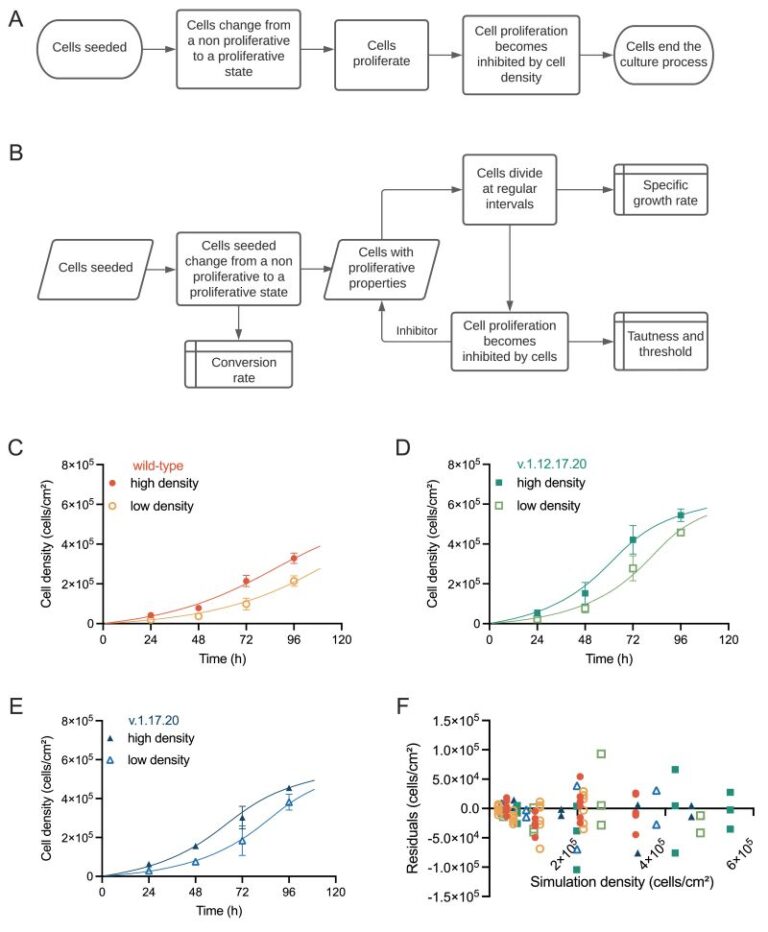
Robert Thomas’s lab at Loughborough University has analyzed growth dynamics between commonly occurring genetically variant hPSCs and their counterpart wild-type cells in culture cells using proprietary computational modelling, allowing the identification of critical process parameters that drive critical quality attributes when genetically variant cells are present within the system This fascinating paper highlights how the system parameters controlling independent growth behavior of wild-type and genetic variant populations are altered when both populations exist within a co-culture environment by introducing an ordinary differential equation (ODE) framework. Findings reveal that variant cells exhibit selective growth and competitive advantage, influencing the behavior of wild-type cells, particularly at higher culture densities. This computational model offers opportunities for defining operational protocols and timely detection of emerging variants, crucial for product release and risk management. It is clear to see the importance as it demonstrates the utility of computational models in understanding complex biological systems and informing manufacturing practices in hPSC-based therapies.
Tan J, Virtue S, Norris DM et al.
Limited oxygen in standard cell culture alters metabolism and function of differentiated cells
From the lab of Daniel J. Fazakerley, University of Cambridge
We all know our stem cell cultures are sensitive and high maintenance, we feed them, keep them warm to make sure we get the most accurate, reproducible data. The composition of their media is important, we need the right growth factors, but this week’s Friday read highlights that their exposure to oxygen can be an important factor in the differentiation and function of cellular models. In this very interesting read from the MRC Institute of Metabolic Science they have investigated the impact of oxygen diffusion and media volume on hypoxia-related transcriptional changes in stem cell cultures. They found that decreasing the media volume decreased lactate production and HIF1α expression and increased the functionality of adipocyte, hiPSC-derived hepatocytes and hiPSC-derived cardiac organoid cultures.
Arboit M, Zorzan I et al.
KLF7 is a general inducer of human pluripotency
From the lab of Elena Carbognin and Graziano Martello, University of Padua
Luo L, Foster NC, Man KL et al.
Hydrostatic pressure promotes chondrogenic differentiation and microvesicle release from human embryonic and bone marrow stem cells
From the lab of Alicia El Haj, University of Birmingham
Williams TL, Colzani MT, Macrae RGC et al.
Human embryonic stem cell-derived cardiomyocyte platform screens inhibitors of SARS-CoV-2 infection
From the lab of Anthony Davenport, University of Cambridge
Ferlazzo GM, Gambetta AM, Amato S et al.
Genome-wide screening in pluripotent cells identifies Mtf1 as a suppressor of mutant huntingtin toxicity
From the lab of Graziano Martello, University of Padova
Tomaz RA, Zacharis ED, Bachinger F et al.
Generation of functional hepatocytes by forward programming with nuclear receptors
From the lab of Ludovic Vallier, University of Cambridge
Rossignoli G, Oberhuemer M, Brun IS et al.
Feeder-free culture of naive human pluripotent stem cells retaining embryonic, extraembryonic and blastoid generation potential
From the lab of Graziano Martello, University of Padua
Stavish D, Price CJ, Gelezauskaite G, Alsehli H et al.
Feeder-free culture of human pluripotent stem cells drives MDM4-mediated gain of chromosome 1q
From the lab of Ivana Barbaric, University of Sheffield
Williams TL, Macrae RGC, Kuc RE, Brown AJH, Maguire JJ, Davenport AP.
Expanding the apelin receptor pharmacological toolbox using novel fluorescent ligands
From the lab of Anthony Davenport, University of Cambridge
Carbognin E, Carlini V, Panariello F et al.
Esrrb guides naive pluripotent cells through the formative transcriptional programme
From the labs of Jamie A. Hackett, European Molecular Biology Laboratory EMBL-Rome, Davide Cacchiarelli, Telethon Institute of Genetics and Medicine and Graziano Martello, University of Padua.
Boutin L, Liu M, Merville JD et al.
EphA2 and phosphoantigen-mediated selective killing of medulloblastoma by γδT cells preserves neuronal and stem cell integrity
From the lab of Margareta T Wilhelm, Karolinska Institute, Stockholm, Sweden
Feofanov M, Daubner GM, Saltalamacchia A et al.
Discovery and optimization of a guanylhydrazone-based small molecule to replace bFGF for cell culture applications
van Bree N, Oppelt AS, Lindström S et al.
Development of an orthotopic medulloblastoma zebrafish model for rapid drug testing
From the lab of Margareta Wilhelm, Karolinska Institutet

The drug discovery process is reliant on appropriate models for high-throughput screening, this can be difficult when complex, heterogeneous diseases are the targeted indication. This publication reports on a fascinating zebrafish model for the study of medulloblastoma, one of the most common malignant brain tumors in children. Introduction of medulloblastoma cells into zebrafish embryos leads to tumor growth in the hindbrain region and the homing of transplanted cells and the aggressiveness of tumor growth were enhanced by pre-culturing cells in a neural stem cell-like medium. This model was then used to successfully assess the effect of anti-cancer drugs on the viability of medulloblastoma cells in this zebrafish embryo model.
Stavish D, Price CJ, Gelezauskaite G et al.
Cytogenetic resource enables mechanistic resolution of changing trends in human pluripotent stem cell aberrations linked to feeder-free culture
From the lab of Ivana Barbaric, University of Sheffield
Used FGF2-G3 (Qk053) at 40 ng/ml and 8 ng/ml for human iPSC culture.
Boikova A, Quaife-Ryan GA, Batho CAP et al.
A transient modified mRNA encoding Myc and Cyclin T1 induces cardiac regeneration and improves cardiac function after myocardial injury
From the labs of Catherine H. Wilson, University of Cambridge and James E. Hudson, QIMR Berghofer Medical Research Institute
Used recombinant activin A (Qk001) at 50 ng/ml and zebrafish FGF-2 (Qk002) at 20 ng/ml for human mesoderm induction.
Ong J, Gibbons G, Lim YS et al.
A clinically defined and xeno-free hydrogel system for regenerative medicine
From the lab of Athina E. Markaki, University of Cambridge
Used recombinant FGF2-G3 (154 aa) protein at 2.25 ng/ml for the culture of cardiac organoids.
Stuart HT et al.
Distinct Molecular Trajectories Converge to Induce Naive Pluripotency
From the lab of José Silva, University of Cambridge
FGF-4
Yamagchi A, Ishikawa K and Akamatsu W
Modeling the pathological brain-gut axis in Parkinson’s disease using human iPSC derived brain-intestinal assembloids
From the lab of Wado Akamatsu, Juntendo University, Tokyo, Japan
FLt3L
Moullet M, Isobe T, Vahidi A et al.
Self-supervised learning for a gene program-centric view of cell states
GDF-15
Ley-Ngardigal S, Claverol S, Sobilo L et al.
Repression of oxidative phosphorylation by NR2F2, MTERF3 and GDF15 in human skin under high-glucose stress
From the lab of Rodrigue Rossignol, University of Bordeaux
Karusheva Y, Ratcliff M, Mörseburg A et al.
The Common H202D Variant in GDF-15 Does Not Affect Its Bioactivity but Can Significantly Interfere with Measurement of Its Circulating Levels
From the lab of Professor Sir Stephen O’Rahilly, University of Cambridge
Fejzo M, Rocha N, Cimino I et al.
GDF15 linked to maternal risk of nausea and vomiting during pregnancy
From the lab of Stephen O’Rahilly, University of Cambridge
The recent publication from Fejzo et al. discusses the role of GDF-15, an intriguing protein acting on the brainstem, in nausea and vomiting during pregnancy, particularly in hyperemesis gravidarum (HG). This study finds that both fetal production of GDF-15 and maternal sensitivity to GDF-15 significantly contribute to the risk of HG with higher levels of GDF15 in maternal blood associated with vomiting during pregnancy and HG. Genetic variants affecting GDF-15 levels influence the risk of HG, with low levels increasing the risk and high levels decreasing it. This was an excellent read which suggests a putative causal role for fetally derived GDF-15 in pregnancy-related nausea and vomiting, with maternal sensitivity influenced by pre-pregnancy exposure and shows great promise for potential avenues of treatment and prevention of HG by blocking GDF-15 action in the pregnant mother.
Fejzo M et al.
Fetally-encoded GDF15 and maternal GDF15 sensitivity are major determinants of nausea and vomiting in human pregnancy
From the lab of Stephen O’Rahilly, University of Cambridge, UK and Nicholas Mancuso, University of Southern California, USA.
Jeromson S, Akcan M, Baranowski B, Arbeau M, Bellucci A, Wright DC.
Daily GDF15 treatment has sex‐specific effects on body weight and food intake and does not enhance the effects of voluntary physical activity in mice
From the lab of David Wright, University of British Columbia
Used human GDF-15 (Qk017) at 0.1 mg/kg for mouse s.c. injection.
Cimino I, Kim H, Tung YCL et al.
Activation of the hypothalamic–pituitary–adrenal axis by exogenous and endogenous GDF15
From the lab of Stephen O’Rahilly, University of Cambridge
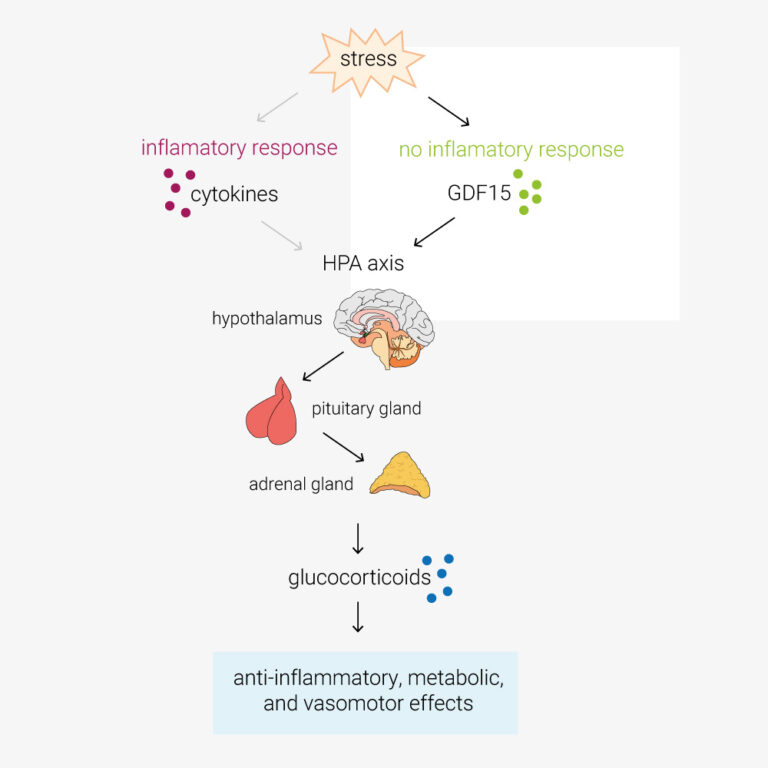
In this paper, Cinimo et al. explore the role of the TGFβ-family protein GDF15 in activation of the hypothalamic–pituitary–adrenal (HPA) axis. During infection, cytokines such as TNFα/β, IL-1 and IL-6, activate the HPA axis. This increases circulating glucocorticoids, which have anti-inflammatory, metabolic, and vasomotor effects. However, O’Rahilly lab have determined that in response to stimuli such as toxins, which don’t provoke an inflammatory response, the primary activator of the HPA axis is GDF15. GDF15 is an intriguing protein also being explored as an anti-obesity therapeutic target, these findings may have a pivotal impact on future clinical study design and open new avenues of investigation. Certainly cool science!
GDNF
Grass T, Cosacak IM, Ordureau A et al.
Isogenic cortical organoids enable precision targeting of APP variant-specific pathways in Alzheimer’s disease
From the lab of Natalia Rodriguez-Muela, DZNE
Used:
Alzheimer’s disease is one of the most prevalent neurodegenerative diseases and the leading cause of dementia, the 7th leading cause of death globally. Currently, there is a lack of suitable and relevant disease models, leading to difficulties in both understanding and treating the disease.
Whilst Alzheimer’s disease primarily affects older adults, early onset familial disease has been shown to be caused by pathogenic mutations in the amyloid precursor protein (APP) gene, these mutations may allow the creation of disease models and therefore a method of testing and developing treatments.
This week’s #Fridayread from the lab of Dr. Natalia Rodriguez-Muela describes the generation of an isogenic panel of hiPSC-derived cortical organoids carrying familial Alzheimer’s disease-associated APP variants or the protective A673T variant. Through the panel of 3D models they identified distinct pathogenic pathways specific to each variant.
Proteomic analysis of the organoids allowed them to determine variant-specific defects which related to proteins dysregulated in post-mortem diseased brains, demonstrating the reliability of their in vitro model. Through this they were able to employ targeted interventions and restore neuronal survival in a variant-specific manner.
This study used Qkine BDNF (Qk050) and GDNF (Qk051) at 20 ng/ml in the differentiation of hiPSCs into cortical spheres.
Thomas J (Thesis)
SMN Protein Heterogeneity Drives Intrinsic Motor Neuron Vulnerability through Survival and Activity Related Pathways
DOI: Thesis
Used BDNF (Qk050) and GDNF (Qk051) at 10 ng/ml for motor neuron differentiation from iPS.
Buchner F, Dokuzluoglu Z, Thomas J et al.
Synchronous 3D patterning of diverse CNS progenitors generates motor neurons of broad axial identity
From the lab of Dr. Natalia Rodriguez-Muela, German Center for Neurodegenerative Diseases e.V. (DZNE)
Page T, Musi CA, Bakker SE et al.
Parkinson’s associated protein DJ-1 regulates intercellular communication via extracellular vesicles in oxidative stress
From the lab of Mariaelena Repici, Aston University
Used:
This publication explores disease mechanisms in an autosomal recessive form of Parkinson’s disease caused by mutations in the DJ-1 protein. Parkinson’s disease is neurodegenerative disorder effecting the central and peripheral nervous system, it is common and effects 1% of people over 60. Parkinson’s is characterized by the loss of dopaminergic neurons and accumulation of Lewy bodies in the substantia nigra area of the brain.
DJ-1 protein, encoded by the Park7 gene, is a multifunctional protein which has been shown to have roles in the regulation of apoptosis ad the immune system and also dopamine synthesis and uptake. This study looks at the role of DJ-1 in extracellular vesicles, which are secreted by brain cells and regulate synaptic function and neural development.
Proteomic analysis of extracellular vesicles from DJ-1 deficient cells showed a distinct protein expression profile. Challenging DJ-1 KO cells with rotenone, a naturally occurring pesticide which induces oxidative stress, showed DJ-1 deficient cells have reduced numbers of extracellular vesicles and increased macrophage migration.
These result indicate DJ-1 regulates the number, cargo and functional effects of extracellular vesicles released from neuronal cells upon oxidative stress, offering new insights into disease progression and novel therapeutic targets.
Agarwal D et al.
Human retinal ganglion cell neurons generated by synchronous BMP inhibition and transcription factor mediated reprogramming
From the lab of Karl Wahlin, University of California San Diego
Used recombinant human BDNF protein (Qk050) at 50 ng/ml and recombinant human GDNF protein (Qk051) at 10 ng/ml for differentiation of PSCs in retinal ganglion cell induced neurons.
Agarwal D & Wahlin K
Differentiation of RGC Induced Neurons (RGC-iNs)
From the lab of Karl Wahlin, University of California San Diego
Gremlin
Sato N, Rosa VS, Makhlouf A et al.
Basal delamination during mouse gastrulation primes pluripotent cells for differentiation
From the lab of Marta Shahbazi, MRC Laboratory of Molecular Biology, Cambridge
Used recombinant human gremlin 1 (Qk015) at 450 ng/ml for culture of mouse epiblast stem cells (EpiSCs).
HGF
Iyer DP, Khoei HH, van der Weijden VA et al.
mTOR activity paces human blastocyst stage developmental progression
From the lab of Dr. Aydan Bulut-Karslıoğlu, Max Planck Institute for Molecular Genetics
IGF
Skowronska-Krawczyk et al.
Age-Driven Lipid Remodeling Activates Lysosome-Mediated Plasma Membrane Repair
From the lab of Lauren Albrecht, University of California, Los Angeles
Leber R, Rosa J, Laizé V et al.
Towards cost-effective and sustainable media formulations for cellular agriculture
From the lab of Aleksandra Fuchs, Austrian Centre of Industrial Biotechnology
Mulholland KE, Bourguet M, Cheng N et al.
Pervanadate-induced oxidation relieves autoinhibition of SRC protein tyrosine kinase
IL-34
Song C, Chen X, Ji R at al.
Identification of secondary microglial formation centers in the human fetal brain
From the lab of Hexige Saiyin, Fudan University, Shanghai, China.
KGF (FGF-7)
Fiancette RB, Huang J, Stephens C et al.
High-purity stem cell-derived β-cells recapitulate key transcriptional and functional features of human islets
From the lab of Dr İldem Akerman, University of Birmingham, UK
Darrigrand J-F, Isaacson A and Spagnoli FM
Generation of human iPSC-derived pancreatic organoids to study pancreas development and disease
Kits
Fischer Y, Mosher J, Barbaric I et al.
Improving rigor and reproducibility through implementation of the ISSCR standards for human stem cell use in research
Review from Yvonne Fischer, Jack Mosher, Ivana Barbaric, Claudia Spits, and Martin F. Pera
LIF
Stuart HT, Costantini E, Wang J at al.
Minimal essential requirements for neural tube self-organisation
Paloviita P, Hautala E, Granskog R et al.
Hsa-miR-92a-3p regulates cell-cycle and signaling programs during human extra-embryonic lineage commitment
From the lab of Sanna Vuoristo, University of Helsinki and Helsinki University Hospital, Finland
Olanipekun JT, Edginton-White B, McQueen C, Geoffrey Brown G and William E.B. Johnson WEB
Retinoic Acid Receptor Gamma Activity Plays a Critical Role in Regulating Early Mouse Gastruloid Development
From the labs of Geoffrey Brown, University of Birmingham, UK and Eustace Johnson, University of Chester, UK.
Artioli A, Gasparotto M, Rossetti AC et al.
A defined 2D system for generating and expanding human basal radial glia from iPSCs
From the lab of Julia Ladewig, Central Institute of Mental Health (ZI), Heidelberg University/Medical Faculty Mannheim, Germany
Zhu K, Li W, Liu J et al.
Functional landscape of non-canonical open reading frames in coordinating cell fate
From the lab of Jin Chen, The University of Texas Southwestern Medical Center, Dallas, TX, USA
Rossignoli G, Oberhuemer M, Brun IS et al.
Serum coating enables feeder-free culture of naive human pluripotent stem cells preserving developmental potential
From the labs of Jan Jakub Żylicz (Novo Nordisk Foundation Center for Stem Cell Medicine, University of Copenhagen, Denmark), Martin Leeb (Max Perutz Labs, University of Vienna, Austria) and Graziano Martello (University of Padua, Italy)
Used:
This publication describes a feeder cell-free culture system based on serum coating that supports long-term maintenance of naive human pluripotent stem cells (hPSCs).
Naive hPSC represent a preimplantation epiblast state able to efficiently differentiate into embryonic and extraembryonic pre-implantation lineages. Their maintenance routinely relies on co-culture with mouse embryonic fibroblast (MEFs) as feeder cells, a method prone to variability.
In a collaborative effort, five independent laboratories tested thirty serum batches for the culture of eight naïve hPSCs lines. Cells cultured on serum coating displayed growth kinetics, clonogenic capacity, mutation rates, and global gene expression profiles comparable to MEF-based cultures. Naive hPSCs efficiently underwent germ layer specification, retained trophectoderm competence, and generated blastoids with efficiency similar to MEF-based cultures.
This study showed that serum coating provides a scalable, cost-effective, and robust alternative to feeder-based systems, facilitating larger-scale applications of naive hPSCs and enabling more reproducible mechanistic studies.
This study used Qkine animal origin-free proteins:
Wang X, Fu, H, Sun Q et al.
Asymmetric Division in a Two-Cell-Like State Rejuvenates Embryonic Stem Cells
From the lab of Mingwei Min, Guangzhou Medical University, China
Chen C, Wu J, Wang X et al.
Signaling reprogramming via Stat3 activation unravels high-fidelity human post-implantation embryo modeling
From the lab of José Silva, Guangzhou National Laboratory, Guangdong, China
Li H, Guan W, Huang J et al.
A complete model of mouse embryogenesis through organogenesis enabled by chemically induced embryo founder cells
From the lab of José Silva, Guangzhou National Laboratory, Guangzhou Medical University, China
Trupej A, Bergant V, Novljan J et al.
HCR-Proxy resolves site-specific proximal RNA proteomes at subcompartmental nanoscale resolution
From the lab of Miha Modic, Karlsruhe Institute of Technology (KIT)
Van Nerum, K., Wenzel, A., Argemi-Muntadas, L. et al.
α-Ketoglutarate promotes trophectoderm induction and maturation from naive human embryonic stem cells
From the lab of Jan Jakub Zylicz, University of Copenhagen
Balayo T, Lunn S, Pascual-Mas P et al.
N2B27 media formulations influence gastruloid development
From the lab of David A Turner, University of Liverpool
Huang, T et al.
Inhibition of PRC2 enables self-renewal of blastoid-competent naive pluripotent stem cells from chimpanzee
From the lab of Hideki Masaki, Institute of Science, Tokyo
Dinarello A, Betto RM, Diamante L et al.
STAT3 and HIF1α cooperatively mediate the transcriptional and physiological responses to hypoxia
From the lab of Graziano Martello and Francesco Argenton, University of Padova
Guo M, Wu J, Chen C et al.
Self-renewing human naïve pluripotent stem cells dedifferentiate in 3D culture and form blastoids spontaneously
From the lab of José Silva, Guangzhou Laboratory
A huge challenge in understanding human early embryo cell fate is due to limited access and ethical concerns. Recent research, however, from José C. R. Silva’s lab at Guangzhou National Laboratory, drawing from single-cell sequencing, suggests a conserved lineage specification process between human and mouse embryos. Blastoids, emerging models for early embryo development, generated solely from hnPSCs, offer insights into blastocyst formation without altering culture conditions. Self-renewing human naïve pluripotent stem cells (hnPSCs) spontaneously form blastoids in 3D culture, mimicking early human blastocysts. This process, mediated by the GSK3 inhibitor IM-12 in 5iLAF medium, involves upregulation of oxidative phosphorylation genes. hnPSCs dedifferentiate into E5 embryo-like intermediates, expressing SOX2/OCT4 and GATA6, which specify trophoblast fate by day 3, coinciding with blastoid formation. This was a fantastic paper to read as it is clear how this spontaneous blastoid formation highlights the importance of culture conditions and provides a new platform to study human embryo development in vitro, potentially reshaping our understanding of hnPSCs and embryo development.
Li H, Chang L, Huang J and Silva JCR.
Protocol for generating mouse morula-like cells resembling 8- to 16-cell stage embryo cells
From the lab of José Silva, Guangzhou Laboratory
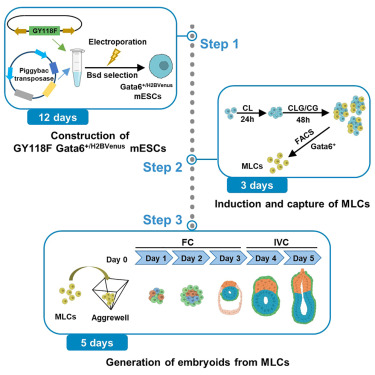
Generating cell types with properties of embryo cells with full developmental potential is of great biological importance. This paper describe steps for induction and isolation of MLCs by sorting. They explained the procedures for segregating MLCs into blastocyst cell fates and how to create embryo-like structures from them. This system provides a valuable stem-cell-based embryo model to study early embryo development.
Rosa VS, Sato N and Shahbazi MN et al.
Protocol for generating a 3D culture of epiblast stem cells
From the lab of Dr Marta Shahbazi, MRC Laboratory of Molecular Biology, Cambridge
Drozd AM, Mariani L, Guo X, Goitea V, Menezes NA and Ferretti E.
Progesterone Receptor Modulates Extraembryonic Mesoderm and Cardiac progenitor Specification during Mouse Gastrulation
Krammer T and Tanaka EM
Neural tube organoid generation: a robust and reproducible protocol from single mouse embryonic stem cells
From the lab of Elly M. Tanaka, Francis Crick Institute and the Research Institute of Molecular Pathology (IMP) of Vienna
Hennessy MJ, Fulton T, Turner DA and Steventon B
Negative feedback on Retinoic Acid by Brachyury guides gastruloid symmetry-breaking
From the labs of David A. Turner, University of Liverpool and Ben Steventon, University of Cambridge
Borkowska M, Leitch HG.
Mouse Primordial Germ Cells: In Vitro Culture and Conversion to Pluripotent Stem Cell Lines
From the lab of Harry Leitch, Imperial College London
Krammer T, Stuart HT, Gromberg E et al.
Mouse neural tube organoids self-organize floorplate through BMP-mediated cluster competition
From the labs of James Briscoe and Elly M. Tanaka, Francis Crick Institute and the Research Institute of Molecular Pathology (IMP) of Vienna

This paper gives us insight into the complex regulatory interactions between BMP-4 and noggin during neural tube development. Neural tube organoids from embryonic stem cells will organize into floorplates, without the usual inducers present in the developing embryo. They used mouse neural tube organoids and by identifying the floorplate marker FOXA2 they showed the importance of regulation of BMP-4 signaling through noggin. Noggin mutation reduced neural tube organization and floorplate formation in vitro and in vivo.
Arboit M, Zorzan I et al.
KLF7 is a general inducer of human pluripotency
From the lab of Elena Carbognin and Graziano Martello, University of Padua
Klobučar T, Novljan J, Iosub IA at el.
Integrative profiling of condensation-prone RNAs during early development
From the lab of Miha Modic, National Institute of Chemistry, Ljubljana, Slovenia
Ferlazzo GM, Gambetta AM, Amato S et al.
Genome-wide screening in pluripotent cells identifies Mtf1 as a suppressor of mutant huntingtin toxicity
From the lab of Graziano Martello, University of Padova
Heidari Khoei H, Javali A, Kagawa H et al.
Generating human blastoids modeling blastocyst-stage embryos and implantation
From the lab of Nicolas Rivron, IMBA, Austrian Academy of Sciences
Rossignoli G, Oberhuemer M, Brun IS et al.
Feeder-free culture of naive human pluripotent stem cells retaining embryonic, extraembryonic and blastoid generation potential
From the lab of Graziano Martello, University of Padua
Carbognin E, Carlini V, Panariello F et al.
Esrrb guides naive pluripotent cells through the formative transcriptional programme
From the labs of Jamie A. Hackett, European Molecular Biology Laboratory EMBL-Rome, Davide Cacchiarelli, Telethon Institute of Genetics and Medicine and Graziano Martello, University of Padua.
Mulas C, Stammers M, Salomaa SI, Heinzen C, Suter DM, Smith A, Chalut KJ.
ERK signalling eliminates Nanog and maintains Oct4 to drive the formative pluripotency transition
From the labs of Austin Smith, University of Cambridge and Kevin J. Chalut, University of Exeter
Ball STM, Hennessy MJ, Tan Y et al.
Domain Specific AI Segmentation of IMPDH2 Rod/Ring Structures in Mouse Embryonic Stem Cells
From the lab of David Turner, University of Liverpool, UK
Used mouse LIF protein (Qk018) for mouse embryonic stem cell culture.
Li H, Huang J, Guan W et al.
Chemically induced cell plasticity enables the generation of high-fidelity embryo model
From the lab of José Silva, Guangzhou Laboratory
Used recombinant mouse LIF (Qk018) at 20 ng/ml for mouse embryonic stem cell culture.
Andrés-San Román JA, Gordillo-Vázquez C, Franco-Barranco D et al.
CartoCell, a high-content pipeline for 3D image analysis, unveils cell morphology patterns in epithelia
From the lab of Luis M. Escudero, University of Sevilla
Used recombinant mouse LIF (Qk018) at 10 ng/ml for mouse embryonic stem cell culture.
Frenster JD, Babin S, Casani-Galdon P et al.
Mosaic gastruloids reveal a temporal restriction for developmental cell competition
From the lab of A. Martinez Arias, University Pompeu Fabra
Used recombinant human activin A PLUS™ protein (Qk005) at 25 ng/ml or 100 ng/ml for gastruloid culture and recombinant mouse LIF protein (Qk018) at 10 ng/ml for mouse embryonic stem cell culture.
Stuart HT et al.
Distinct Molecular Trajectories Converge to Induce Naive Pluripotency
From the lab of José Silva, University of Cambridge
Noggin
Hsu J, Song H, Ogawa S et al.
CA19-9 induces microenvironment remodeling in pancreatic ductal adenocarcinoma
From the lab of Dannielle Engle, Salk Institute for Biological Studies, La Jolla, California, USA
Takase Y, Murata Y, Namba K and Takahashi T
A region-specific murine intestinal monolayer platform for assessing iron form–dependent transepithelial transport
From the lab of Toshio Takahashi, Bioorganic Research Institute, Kyoto, Japan
Tran D, Tolley C, Morris T et al.
Recapitulating whipworm development in vitro using caecaloids
From the lab of Dr Maria Duque-Correa, University of Cambridge
Tanaka E, Kishikawa T, Miyakawa Y et al.
Modeling intestinal epithelial function and polarity using human iPSCs under Air–Liquid interface culture
From the lab of Mitsuhiro Fujishiro, University of Tokyo, Japan
Lumibao JC, Okhovat SR, Peck KL et al.
The effect of extracellular matrix on the precision medicine utility of pancreatic cancer patient-derived organoids
From the lab of Dannielle Engle, Salk Institute for Biological Studies

A promising application of organoids, in this case patient-derived organoids (PDOs), is to profile the therapeutic sensitivity of tumors to inform clinical decisions and to stratify patients in clinical trials more effectively. Recently there have been positive results from clinical trials giving a glimmer of the potential impact of these technologies. However, a concern is how reproducible these technologies are, particularly as most still rely on animal-derived basement membrane extracts, where batch variation is a sector-wide concern. This recent publication shows consistent drug responses are observed when different sources of commercial BME were compared. This is reassuring and speaks to how robust these approaches can be.
Bergmann S, Penfold CA, Slatery E et al.
Spatial profiling of early primate gastrulation in utero
From the lab of Thorsten E. Boroviak, University of Cambridge
Osorio-Vasquez V, Lumibao JC, Peck KL et al.
Chronic pancreatitis patient-derived organoids reveal new paths to precision therapeutics
From the lab of Dannielle D. Engle, Salk Institute for Biological Studies
Agarwal R, Dittmar T, Beer HD et al.
Human epidermis organotypic cultures, a reproducible system recapitulating the epidermis in vitro
From the labs of Emmanuel Contassot and Alexander A. Navarini, University of Basel
NRG-1
Hernanz M, Matas D, Mirasierra M et al.
Intramuscular Transplantation of Human Pluripotent Stem Cell-derived Pancreatic Endocrine Cells in Mice
DOI: DOI: 10.3791/69562
From the lab of Alberto Bartolomé, Biomedical Research Institute, Madrid
Bottini S (Thesis)
Animal proteins from stem cells as an alternative to reduce the ecological and climate impact of animal farming
DOI: Thesis
Used:
- Recombinant bovine/porcine FGF-2 (145 aa) protein (Qk040-FG)
- Recombinant bovine/porcine FGF-2 (145 aa) protein (Qk040)
- Recombinant human TGF-β1 PLUS™ protein (Qk010-CTG)
- Recombinant human TGF-β1 PLUS™ protein (Qk010)
- Recombinant FGF2-G3 (145 aa) protein (Qk052)
- Recombinant human TGF-β3 protein (Qk054-CTG)
- Recombinant human TGF-β3 protein (Qk054)
- Recombinant human NRG-1 protein (Qk045)
- Recombinant human activin A protein (Qk001-CTG)
- Recombinant human activin A protein (Qk001)
- Recombinant human BMP-4 protein (Qk038)
Truszkowski L, Bottini S, Bianchi S et al.
Refined and benchmarked homemade media for cost-effective, weekend-free human pluripotent stem cell culture
From the lab of Alessandro Bertero, University of Turin in collaboration with Qkine
Used:
- Recombinant human activin A protein (Qk001)
- Recombinant human FGF-2 (145 aa) protein (Qk025)
- Recombinant human FGF-2 (154 aa) protein (Qk027)
- Recombinant human BMP-4 protein (Qk038)
- Recombinant human NRG-1 protein (Qk045)
- Recombinant FGF2-G3 (145 aa) protein (Qk052)
- Recombinant FGF2-G3 (154 aa) protein (Qk053)
- Recombinant human TGF-β3 protein (Qk054)
- Recombinant human TGF-β1 PLUS™ protein (Qk010)
- Thermostable FGF-2 discovery kit (Qk502)
Cell therapy is becoming a possibility for many previously untreatable conditions, and it should be accessible to everyone. Creating a cost-effective, reliable and reproducible way of culturing human induced pluripotent stem cells (hiPSCs) in a range of research labs, and allowing large scale culture for gene-editing purposes takes us one step closer to this.
Using high potency thermostable Qkine 145 amino acid FGF-G3 reduce FGF-2 use 8-fold and for weekend-free culture reduced media use by 57%. This makes hiPSCs a more accessible model for many labs doing basic and translational research.
NT-3
Sozza F, Romano A, D’Elia N et al.
Combinatorial and Inducible CRISPRa/i Enables Canalized hiPSC Forward Programming and Iterative Refinement via Single-Cell Genomics
From the lab of Alessandro Bertero, Department of Molecular Biotechnology and Health Sciences, University of Turin, Italy
Used:
- Recombinant human NT-3 protein (Qk058)
- Recombinant human activin A protein (Qk001-CTG)
- Recombinant human activin A protein (Qk001)
- Recombinant human BDNF protein (Qk050-CTG)
- Recombinant human BDNF protein (Qk050)
- Recombinant human FGF-2 (145 aa) protein (Qk025-CTG)
- Recombinant human FGF-2 (145 aa) protein (Qk025)
- Recombinant human BMP-4 protein (Qk038)
Liu Y, Li X, Xu H et al.
Spinal cord stimulation induces Neurotrophin-3 to improve diabetic foot disease
PDGF
Mulholland KE, Bourguet M, Cheng N et al.
Pervanadate-induced oxidation relieves autoinhibition of SRC protein tyrosine kinase
Qkine
Fischer Y, Mosher J, Barbaric I et al.
Improving rigor and reproducibility through implementation of the ISSCR standards for human stem cell use in research
Review from Yvonne Fischer, Jack Mosher, Ivana Barbaric, Claudia Spits, and Martin F. Pera
Wonfor RE, Ong KJ, Ng W et al.
Standardising Culture Medium Safety Testing for Cultivated Meat: Outputs from a Workshop and Case Study
Report collated by Cai Linton, Multus Bio
Barber L, Spicer C.
The effect of pyridinecarboxaldehyde functionalisation on reactivity and N-terminal protein modification
From the lab of Christopher Spicer, University of York
Karusheva Y, Ratcliff M, Mörseburg A et al.
The Common H202D Variant in GDF-15 Does Not Affect Its Bioactivity but Can Significantly Interfere with Measurement of Its Circulating Levels
From the lab of Professor Sir Stephen O’Rahilly, University of Cambridge
Barber LJ, Yates NDJ, Fascione MA et al.
Selectivity and stability of N-terminal targeting protein modification chemistries
Venkatesan M, Semper C, Skrivergaard S et al.
Recombinant production of growth factors for application in cell culture
From the lab of Alexi Savchenko, University of Toronto
R-spondin
Hsu J, Song H, Ogawa S et al.
CA19-9 induces microenvironment remodeling in pancreatic ductal adenocarcinoma
From the lab of Dannielle Engle, Salk Institute for Biological Studies, La Jolla, California, USA
Takase Y, Murata Y, Namba K and Takahashi T
A region-specific murine intestinal monolayer platform for assessing iron form–dependent transepithelial transport
From the lab of Toshio Takahashi, Bioorganic Research Institute, Kyoto, Japan
Park M-S, Gunassekaran GR, Lee S-M et al.
Mesothelin-binding peptide inhibits cell migration and enables targeted delivery of mitochondrial membrane–damaging peptide to pancreatic tumors
From the lab of Byungheon Lee, Kyungpook National University, Republic of Korea
Uefune F and Yui S
A murine small intestinal two-dimensional monolayer platform that induces fate commitment toward a revival stem cell state
From the lab of Shiro Yui, Institute of Science Tokyo, Japan
Tsang CF, Patel H, Kouassi FM et al.
Utility of pancreatic tumor scrapings for organoid development and precision medicine strategies
From the labs of Amber Habowski and David Tuveson, Cold Spring Harbor Laboratory, USA.
Yamagchi A, Ishikawa K and Akamatsu W
Modeling the pathological brain-gut axis in Parkinson’s disease using human iPSC derived brain-intestinal assembloids
From the lab of Wado Akamatsu, Juntendo University, Tokyo, Japan
Sung Y, Yu YC, Lee M et al.
Targeting cancer glutamine dependency with a first-in-class inhibitor of the mitochondrial glutamine transporter SLC1A5_var
From the lab of Jung Min Han at the Yonsei Institute of Pharmaceutical Sciences, Republic of Korea
Used recombinant human R-spondin 1 protein (Qk006) at 50 ng/ml for patient-derived pancreatic tumor organoid culture.
Kang S, Lee MR, Choi W, Kong S-Y and Kim Y-H
Protocol for generation and utilization of patient-derived organoids from multimodal specimen
From the lab of Yun-Hee Kim, Research Institute of National Cancer Center, Republic of Korea
Kirino S, Uefune F, Miyake K et al.
Fetal reversion from diverse lineages sustains the intestinal stem cell pool and confers stress resilience
From the lab of Shiro Yui, Center for stem cell and regenerative medicine, Institute of Biomedical Engineering, Institute of Science Tokyo, Japan
Lee, Mi Rim et al.
Organoid morphology-guided classification for oral cancer reveals prognosis
From the lab of Yun-Hee Kim, Research Institute of National Cancer Center, Republic of Korea
Lumibao JC, Okhovat SR, Peck KL et al.
The effect of extracellular matrix on the precision medicine utility of pancreatic cancer patient-derived organoids
From the lab of Dannielle Engle, Salk Institute for Biological Studies
A promising application of organoids, in this case patient-derived organoids (PDOs), is to profile the therapeutic sensitivity of tumors to inform clinical decisions and to stratify patients in clinical trials more effectively. Recently there have been positive results from clinical trials giving a glimmer of the potential impact of these technologies. However, a concern is how reproducible these technologies are, particularly as most still rely on animal-derived basement membrane extracts, where batch variation is a sector-wide concern. This recent publication shows consistent drug responses are observed when different sources of commercial BME were compared. This is reassuring and speaks to how robust these approaches can be.
Ku B, Eisenbarth D, Baek S et al.
PRMT1 promotes pancreatic cancer development and resistance to chemotherapy
From the lab of Dae-Sik Lim, KAIST, Republic of Korea
Marsee A, Ritchie A, Myszczyszyn A et al.
Mass Generation and Long-term Expansion of Hepatobiliary Organoids from Adult Primary Human Hepatocytes
From the lab of Bart Spee, Hubrecht Institute, The Netherlands
Osorio-Vasquez V, Lumibao JC, Peck KL et al.
Chronic pancreatitis patient-derived organoids reveal new paths to precision therapeutics
From the lab of Dannielle D. Engle, Salk Institute for Biological Studies
Agarwal R, Dittmar T, Beer HD et al.
Human epidermis organotypic cultures, a reproducible system recapitulating the epidermis in vitro
From the labs of Emmanuel Contassot and Alexander A. Navarini, University of Basel
Choi W, Kim YH, Woo SM et al.
Establishment of Patient-Derived Organoids Using Ascitic or Pleural Fluid from Cancer Patients
From the lab of Sun-Young Kong, National Cancer Center, Korea
SCF
Moullet M, Isobe T, Vahidi A et al.
Self-supervised learning for a gene program-centric view of cell states
Azami T, Theeuwes B, Ton M-LN et al.
STAT3 signalling enhances tissue expansion during postimplantation mouse development
From the lab of Dr Jennifer Nichols, University of Cambridge
StemCultures
TCW J, Qian L, Pipalia NH et al.
Cholesterol and matrisome pathways dysregulated in astrocytes and microglia
From the lab of Alison M Goate, Department of Genetics and Genomic Sciences, Icahn School of Medicine at Mount Sinai, New York
Lotz S, Goderie S, Tokas N et al.
Sustained levels of FGF2 maintain undifferentiated stem cell cultures with biweekly feeding
From the lab of Christopher A. Fasano Neural Stem Cell Institute, Rensselaer, New York
Bertucci T, Kakarla S, Winkelman MA et al.
Direct differentiation of human pluripotent stem cells into vascular network along with supporting mural cells
From the lab of Guohao Dai, Northeastern University, Boston, Massachusetts
TGF-β
Hernanz M, Matas D, Mirasierra M et al.
Intramuscular Transplantation of Human Pluripotent Stem Cell-derived Pancreatic Endocrine Cells in Mice
DOI: DOI: 10.3791/69562
From the lab of Alberto Bartolomé, Biomedical Research Institute, Madrid
Bottini S (Thesis)
Animal proteins from stem cells as an alternative to reduce the ecological and climate impact of animal farming
DOI: Thesis
Used:
- Recombinant bovine/porcine FGF-2 (145 aa) protein (Qk040-FG)
- Recombinant bovine/porcine FGF-2 (145 aa) protein (Qk040)
- Recombinant human TGF-β1 PLUS™ protein (Qk010-CTG)
- Recombinant human TGF-β1 PLUS™ protein (Qk010)
- Recombinant FGF2-G3 (145 aa) protein (Qk052)
- Recombinant human TGF-β3 protein (Qk054-CTG)
- Recombinant human TGF-β3 protein (Qk054)
- Recombinant human NRG-1 protein (Qk045)
- Recombinant human activin A protein (Qk001-CTG)
- Recombinant human activin A protein (Qk001)
- Recombinant human BMP-4 protein (Qk038)
Skowronska-Krawczyk et al.
Age-Driven Lipid Remodeling Activates Lysosome-Mediated Plasma Membrane Repair
From the lab of Lauren Albrecht, University of California, Los Angeles
Minerath RA, Kasam RK, Swoboda CO et al.
Cardiomyocyte-expressed TGFβ signals to fibroblasts to program early heart maturation and adult myocyte identity
From the lab of Jeffery Molkentin, Cincinnati Children’s Hospital Medical Center, University of Cincinnati College of Medicine, USA
Rastovic U, Campinoti S, Wei L et al.
Comprehensive analysis of extracellular matrix remodelling via cyclophilin inhibition in human models of alcohol-related liver fibrosis
From the lab of Elena Palma, Foundation for Liver Research and King’s College Hospital, London, UK
Mulholland KE, Bourguet M, Cheng N et al.
Pervanadate-induced oxidation relieves autoinhibition of SRC protein tyrosine kinase
Shin D, Kim CN, Ross J et al.
Thalamocortical organoids enable in vitro modeling of 22q11.2 microdeletion associated with neuropsychiatric disorders
From the lab of Tomasz Nowakowski, University of California, San Francisco

This recent study from David Shin in the lab of Tomasz J. Nowakowski University of California, explores thalamic dysfunction in psychiatric disorders, focusing on the 22q11.2 microdeletion associated with increased risk. They used human pluripotent stem cell-derived organoids to investigate early thalamus development, revealing widespread transcriptional dysregulation and elevated FOXP2 expression in thalamic neurons and glia. From a co-culture model, they found that the microdeletion leads to thalamic axon overgrowth, mediated by FOXP2. These findings suggest dysregulated thalamic development contributes to schizophrenia-related neural phenotypes in 22q11.2 deletion syndrome, offering fascinating insights into the neuropsychiatric disorder’s genetic mechanisms. It will be very interesting to see how human pluripotent stem cell-derived organoids continue to play a fundamental role in psychiatric disorders in the future!
Truszkowski L, Bottini S, Bianchi S et al.
Refined and benchmarked homemade media for cost-effective, weekend-free human pluripotent stem cell culture
From the lab of Alessandro Bertero, University of Turin in collaboration with Qkine
Used:
- Recombinant human activin A protein (Qk001)
- Recombinant human FGF-2 (145 aa) protein (Qk025)
- Recombinant human FGF-2 (154 aa) protein (Qk027)
- Recombinant human BMP-4 protein (Qk038)
- Recombinant human NRG-1 protein (Qk045)
- Recombinant FGF2-G3 (145 aa) protein (Qk052)
- Recombinant FGF2-G3 (154 aa) protein (Qk053)
- Recombinant human TGF-β3 protein (Qk054)
- Recombinant human TGF-β1 PLUS™ protein (Qk010)
- Thermostable FGF-2 discovery kit (Qk502)
Cell therapy is becoming a possibility for many previously untreatable conditions, and it should be accessible to everyone. Creating a cost-effective, reliable and reproducible way of culturing human induced pluripotent stem cells (hiPSCs) in a range of research labs, and allowing large scale culture for gene-editing purposes takes us one step closer to this.
Using high potency thermostable Qkine 145 amino acid FGF-G3 reduce FGF-2 use 8-fold and for weekend-free culture reduced media use by 57%. This makes hiPSCs a more accessible model for many labs doing basic and translational research.
Venkatesan M, Semper C, Skrivergaard S et al.
Recombinant production of growth factors for application in cell culture
From the lab of Alexi Savchenko, University of Toronto
Lyra-Leite DM, Copley RR, Freeman PP et al.
Nutritional requirements of human induced pluripotent stem cells
From the lab of Paul W. Burridge, Northwestern University Feinberg School of Medicine
Beltran-Rendon C, Price CJ, Glen K et al.
Modeling the selective growth advantage of genetically variant human pluripotent stem cells to identify opportunities for manufacturing process control
From the lab of Robert Thomas, Loughborough University
Robert Thomas’s lab at Loughborough University has analyzed growth dynamics between commonly occurring genetically variant hPSCs and their counterpart wild-type cells in culture cells using proprietary computational modelling, allowing the identification of critical process parameters that drive critical quality attributes when genetically variant cells are present within the system This fascinating paper highlights how the system parameters controlling independent growth behavior of wild-type and genetic variant populations are altered when both populations exist within a co-culture environment by introducing an ordinary differential equation (ODE) framework. Findings reveal that variant cells exhibit selective growth and competitive advantage, influencing the behavior of wild-type cells, particularly at higher culture densities. This computational model offers opportunities for defining operational protocols and timely detection of emerging variants, crucial for product release and risk management. It is clear to see the importance as it demonstrates the utility of computational models in understanding complex biological systems and informing manufacturing practices in hPSC-based therapies.
VEGF
Akiyama H, Katayama Y, Shimizu K and Honda H
Engineering a controlled cardiac multilineage co-differentiation process using statistical design of experiments
From the lab of Hiroyuki Honda, Nagoya University, Japan.
Azami T, Theeuwes B, Ton M-LN et al.
STAT3 signalling enhances tissue expansion during postimplantation mouse development
From the lab of Dr Jennifer Nichols, University of Cambridge
Zambaldi V, La D, Chu AE et al.
De novo design of high-affinity protein binders with AlphaProteo
Used human VEGF 165 (Qk048) for x-ray crystallography.