Differentiation of induced pluripotent stem cells (iPSCs) into endoderm
Leighton Sneade
Qkine
The differentiation of feeder-free human induced pluripotent stem cells (iPSCs) into endodermal cells is a complex process involving growth factors like activin A, BMP4, and FGF2. Additionally, factors such as CHIR99021 and the phosphatidylinositol 3-kinase (PI3-kinase) inhibitor LY294002 are required to improve the efficiency of differentiation and stabilize the endodermal fate. This combination allows researchers to reliably generate functional definitive endodermal (DE) cells for various biomedical applications.
In this application note, we outline the method for differentiating iPSCs into definitive endoderm (DE) cells using media containing members of our recombinant FGF2 family, such Qkine FGF2-G3 (Qk053), along with both activin A (Qk001), and BMP4 (Qk038) growth factors. DE marker expression is used to assess successful differentiation via immunocytochemistry.
Introduction
Human induced pluripotent stem cells (iPSCs) are an in vitro model that represent a pivotal breakthrough in regenerative medicine and cellular biology. iPSCs are generated by reprogramming adult somatic cells to a pluripotent state through the introduction of specific transcription factors. Reprogramming iPSCs grants these cells the ability to differentiate into any cell type of the three germ layers: ectoderm, mesoderm, and endoderm. This provides unparalleled potential for disease modeling, drug discovery, and cell-based therapies, all without the ethical concerns associated with using embryonic stem cells [1].
Differentiating iPSCs into endodermal linage cells is particularly important due to the critical roles endodermal derivatives play in human physiology. The endoderm layer forms essential organs like the liver, pancreas, lungs, and the lining of the gastrointestinal tract [2]. Understanding the differentiation process of iPSCs into endodermal cells offers promise for developing treatments for a variety of conditions, including liver diseases, diabetes, and cystic fibrosis, as well as for generating organoids for research and transplantation purposes.
The differentiation process of iPSCs into endodermal cells typically involves mimicking the stages of embryonic development in vitro. During embryogenesis, endoderm formation is initiated by a series of signaling pathways that include nodal, Wnt, and activin/nodal, which are essential for mesendoderm specification and subsequent DE development [3,4]. To mimic these conditions in a controlled laboratory environment, researchers utilize specific growth factors and small molecules to guide iPSCs through comparable developmental signals.
The successful differentiation of iPSCs into endodermal cells comes with its challenges. Variability within iPSC lines, differentiation efficiencies, and the potential for incomplete or mixed lineage differentiation are significant obstacles that researchers continue to tackle. Evaluating success involves examining the expression of markers such as SRY-box transcription factor 17 (SOX17) and transcription factor GATA4 [5].
References
[1] Varum, S. et al. Energy Metabolism in Human pluripotent stem cells and their differentiated counterparts. PLoS ONE. 2011;6(6):e20914. doi: 10.1371/journal.pone.0020914
[2] Zorn, A. M. et al. Vertebrate Endoderm Development and Organ Formation. Annual review of cell and development biology 2009:25:221-51. doi: 10.1146/annurev.cellbio.042308.113344
[3] Fang, Y. et al. Metabolic and Epigenetic Regulation of Endoderm Differentiation. Trends in Cell biology 2022 Feb;32(2):151-164. doi: 10.1016/j.tcb.2021.09.002
[4] Ikonomou, L. et al. Derivation of endodermal progenitors from pluripotent stem cells. Journal of Cellular Physiology. 2015 Feb;230(2):246-58 doi: 10.1002/jcp.24771
[5] McLean, A. B. et al. Activin A Efficiently Specifies Definitive Endoderm from Human Embryonic Stem Cells Only When Phosphatidylinositol 3-Kinase Signaling Is Suppressed. Stem Cells. Jan;25(1):29-38. doi: 10.1634/stemcells.2006-0219
Materials and Methods
Cell culture and maintenance
iPSCs were passaged twice per week using 0.5 mM EDTA for detachment and seeded in vitronectin (Qk120) (5 µg/ml) coated 6-well plates using a 1:6 split ratio and cultured in an E8-like media. The day after passage, spent media was removed to be replaced with 5 ml of E8-like media allowing the cells to follow a weekend-free media change pattern. For further information on this process, please see our guide to Weekend-free human induced pluripotent stem cell culture using thermostable FGF-2 (bFGF) from Qkine, together with our animal origin-free TGF-β1 and vitronectin, for improved colony homogeneity.
iPSC differentiation into definitive endoderm
The DE differentiation workflow schematic (Figure 1) outlines the steps for the differentiation of iPSCs into DE and assessing differentiation by testing DE expression markers.

Figure 1. Schedule for endoderm differentiation and evaluation testing
iPSCs were detached using AccutaseTM and seeded at 1,000 cells / well in a vitronectin (Qk120) (5 µg/ml) coated 96-well plate in E8-like media containing ROCK inhibitor (Y-27632, 10 µM). The following day, Day 1, cells were fed with freshly prepared DE 1 medium (Table 1). On Day 2, cells were fed with freshly prepared DE 2 medium (Table 1) and on Day 3, cells were fed with freshly prepared DE 3 medium (Table 1).
| Media and supplements | DE 1 medium | DE 2 medium | DE 3 medium |
|---|---|---|---|
| CDM-PVA (Table 2) | Base | Base | – |
| RPMI 1640 (Fisher Scientific 11569726) | – | – | Base |
| Recombinant Activin A (Qk001) | 100 ng/ml | 100 ng/ml | 100 ng/ml |
| Recombinant FGF2 (Qk053) | 80 ng/ml | 80 ng/ml | 80 ng/ml |
| Recombinant BMP-4 (Qk038) | 10 ng/ml | 10 ng/ml | – |
| Ly294002 (Promega V1201) | 10 µM | 10 µM | – |
| CHIR99021 (Stratech S2924-SEL) | 3 µM | – | – |
| B27 supplement (Thermo Scientific 17504044) | – | – | 1x |
| Non-Essential Amino Acid (100x) (Thermo Scientific 11140035) | – | – | 1x |
Table 1. DE medium construction components. Components added aseptically before use.
Data shown from FGF2-G3 (Qk053), this protocol has also been successfully completed with recombinant zebrafish FGF-2 (bFGF) protein (Qk002), human FGF-2 145 aa (Qk025), human FGF-2 154 aa (Qk027), bovine/porcine FGF-2 145aa (Qk040), mouse FGF-2 (Qk042), FGF2-G3 145 aa (Qk052) and bovine/porcine FGF-2 154 aa (Qk056).
| CDM-PVA medium | |
|---|---|
| F-12 Nutrient Mix (Thermo Scientific 31765027) | Half of base media |
| IMDM (Fisher Scientific 11510596) | Half of base media |
| 5% PVA solution [5 g PVA (Merck P8136) in 100 ml water from embryo transfer (Merck W1503)] | 0.1% |
| CD concentrated Lipids (Thermo Scientific 11905031) | 1% |
| Transferrin (Merck T1147) | 15 µg/ml |
| 1-Thioglycerol (Merck M6145) | 0.5 mM |
Table 2. CDM-PVA medium construction components. Components added aseptically and media filtered before use.
Immunocytochemistry
On Day 4, cells were fixed with 4% paraformaldehyde, blocked and permeabilized with 10% donkey serum diluted in 0.1% Triton X-100. Specific antibodies for endodermal markers GATA4 and SOX17 were applied for immunostaining overnight at 4°C. iPSCs were then washed and incubated with the secondary antibodies Donkey anti-Mouse AlexaFluorTM 488 or Donkey anti-Goat AlexaFluorTM 488 together with Hoechst 33258, followed by imaging in phosphate-buffered saline.
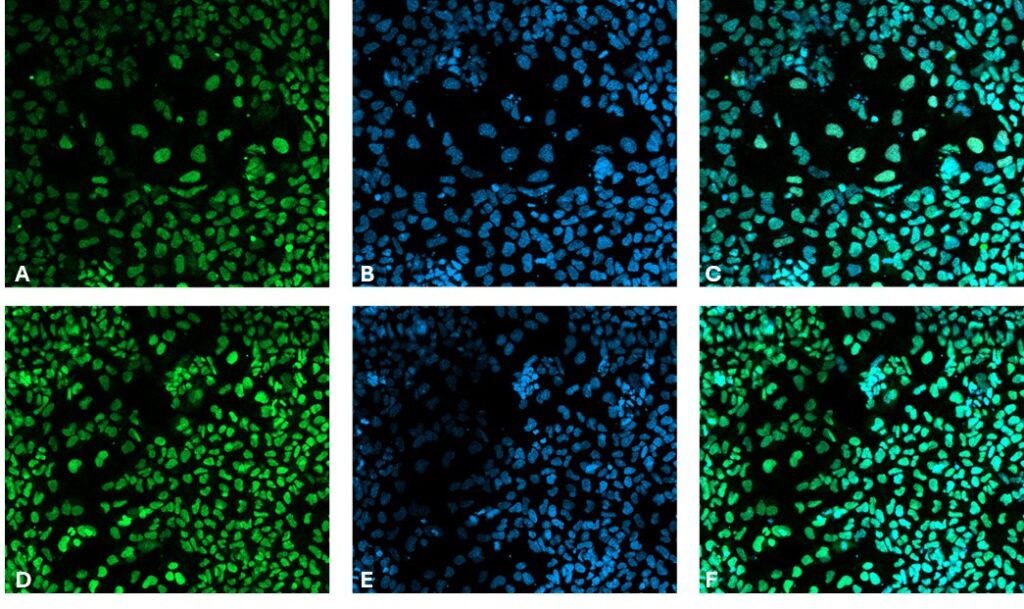
Figure 2. Immunocytochemistry of endoderm markers in differentiated iPSC. SRY-box transcription factor 17 (SOX17) [Green, A], Hoechst 33258 [Blue, B], combined SOX17 and Hoechst [C] and transcription factor GATA4 [Green, D], Hoechst33258 [Blue, E], combined GATA4 and Hoechst [F]. Images were acquired using the tile functionality of the Zeiss LSM 980 with Airyscan2 at 10x magnification.
Conclusion
The differentiation of iPSCs into DE cells is an advancing and highly promising field with considerable therapeutic potential. By leveraging our understanding of embryonic development and refining differentiation protocols, researchers can reliably generate functional DE cells for a variety of biomedical applications.
The data presented in this application note demonstrates that using Qkine FGF-2, activin A, and BMP-4 supports the differentiation of iPSCs into DE cells.
Further Information
Qkine growth factors are manufactured to the highest of quality standards and are free from animal-derived contaminants, delivering low endotoxicity and high purity. At Qkine, we are committed to raising the standards of growth factors, cytokines and related proteins to better support long-term and complex neural stem cell culture. We are a science-led team, please reach out with any questions or requests to support@qkine.com.
All our recombinant proteins are animal-free and come with Bioactivity. Guaranteed.
Contact us
Our science team is here to help, please contact us if you have any questions.